1. Введення
Діабет — це захворювання, у якому порушення регуляції передачі сигналів інсуліну призводить до инсулинорезистентности
1 ]. Клінічні дослідження показали, що глікемічна регуляція та порушення виділення інсуліну негативно впливають на функцію мозку.
2 ]. Крім того, кілька досліджень показали, що діабет є фактором ризику когнітивних захворювань, таких як судинна деменція та хвороба Альцгеймера.
3 ]. Відповідно, літні пацієнти з діабетом 2 типу (ЦД2) отримують клінічні рекомендації щодо позитивного впливу вправ на підвищення чутливості до інсуліну.
4 ]. Однак кілька всеосяжних посібників продемонстрували зв'язок між когнітивною дисфункцією та діабетом.
Ендоплазматичний ретикулум (ЕР) відіграє важливу роль у синтезі білків, а також у згортанні та процесингу секреторних та трансмембранних білків [
5 ]. Коли неправильно згорнуті або розгорнуті білки накопичуються в ER через генетичні фактори або фактори навколишнього середовища, виникає стрес ER, який опосередковується трьома трансмембранними білками ER, IRE1, PERK і ATF6, викликаючи відповідь розгорнутих білків (UPR) [
6 ].
Оскільки стрес ER може активувати клітинні запальні шляхи, які порушують клітинні функції та призводять до метаболічних порушень, існуючі дані свідчать про те, що стрес ER є ключовою ланкою між T2DM, ожирінням та інсулінорезистентністю.
7 ,
8 ]. Відомо, що вплив високих рівнів глюкози може спричинити стрес ER за рахунок генерації вільних радикалів, аберантного глікозилювання білка та збільшення мембранного та білкового обміну, що призводить до посилення експресії білка гомології C/EBP (CHOP) (відомий медіатор стресу ER-інду).
Діабет також пов'язаний з церебральною атрофією, неврологічними порушеннями, зниженням когнітивних функцій та психомоторною дисфункцією, що називається діабетичною енцефалопатією (ДЕП) [
9 ]. Зростання кількості даних вказує на те, що окисний стрес, хронічне запалення, мітохондріальна дисфункція, порушення передачі сигналів інсуліну та стрес ER сприяють розвитку DEP. Однак мало досліджень вивчали вплив кишкового стресу ЕР.
Повідомлялося, що на початку стресу ER підвищена експресія GRP78 у головному мозку щура запобігала індукованому CHOP апоптозу, але коли стрес ER був стійким, рівні CHOP помітно підвищувалися в гіпокампі щурів з діабетом [
10 ]. Це передбачає, що апоптоз, опосередкований стресом CHOP-ER, може бути залучений до індукованих гіперглікемією синапсів гіпокампу та нейрональних порушень і, таким чином, сприяти діабетичним когнітивним порушенням.
Куркумін — це природна сполука з відомими антиоксидантними та протизапальними властивостями [
11 ,
12 ]. Хоча було показано, що він викликає апоптоз ракових клітин, його вплив на запальну реакцію при ЦД2 недостатньо вивчений. Дані, що накопичуються, вказують на те, що слабке запалення, окисний стрес і стрес ER сприяють резистентності до інсуліну при ЦД2. Отже, схоже, що пригнічення стресу та запалення ER може покращити когнітивні функції при ЦД2.
13 ]. Однак не було встановлено, чи вправи в поєднанні з додаванням куркуміну поліпшити когнітивну дисфункцію, пов'язану з діабетом, шляхом регулювання стресової реакції ER.
У цьому дослідженні ми досліджували, чи розрізняються когнітивні функції у щура Otsuka Long-Evans Tokushima Fatty (OLETF) (щур з діабетом) і у Long-Evans Tokushima Otsuka (LETO; щур з контролю), і чи існує комбінація вправи та добавка курей когнітивну дисфункцію, пов'язану з діабетом. Крім того, ми виміряли зміни у рівнях цитокінів запальної відповіді та CHOP, спричинені комбінацією вправ та добавок куркуміну у моделі спонтанного діабету на щурах.
2. Матеріали та методи
2.1. Тварини та експериментальний дизайн
Як контроль використовували щурів LETO (контрольні самці LETO;
n = 10), а як модель СД2 використовували щурів OLETF, гіперфагічний фенотип з мутаціями рецептора холецистокініна-1 [
14 ]. Самців щурів LETO та OLETF містили по два у клітці при температурі 20 ± 2,5°C та 12:12 годинному циклі світло-темрява у світлому приміщенні, постачали достатнє харчування та воду, а також контролювали масу тіла та споживання їжі. Пацюків годували або стандартною їжею (SC; 10 ккал% жиру, дієта D12492 відповідає 7% сахарози, дослідницька дієта, США; контроль OLETF), або стандартною дієтою (SC), що містить куркумін (дієта 5 г/кг, Sigma, Каталожний номер C1386). Пацюки OLETF були випадково розділені на три групи (контрольна група OLETF;
n = 10), діабетична група, яка виконувала вправи (група OLETF + EX;
n= 10), або група діабетиків, які тренувалися плюс куркумін (група OLETF + EX + куркумін);
n = 10) у віці 22 тижнів. Самці щурів LETO були зіставлені за віком. Усі результати експериментів на тваринах були схвалені Комітетом з використання та догляду за тваринами в Університеті Ханнам (HNU2016-16).
2.2. Протокол вправ
Щурів у групі OLETF+EX навчали підніматися вертикальними сходами довжиною 1 м (нахил 85°) з вантажами, прикріпленими до їх хвостів. Протягом першого тижня щурів привчали забиратися на верхню клітку з вантажем або без нього. Тренувальні заняття з другого тижня розпочинали з вагою 50% від маси тіла. Щури почали підніматися з нижньої частини сходів і були змушені піднятися нагору. Коли вони досягли вершини, їм дали 2-хвилинний відпочинок перед наступним випробуванням. Наступні випробування проводили знизу сходів, і при кожному випробуванні до попередньої ваги додавали 20 г. Якщо щур міг піднятися 10 разів зі збільшенням ваги, тренування вважалося завершеним. У разі невдачі при підйомі щур був змушений виконати 10 спроб з останньою успішною вагою без подальшого збільшення ваги [
15 ].
2.3. Водний лабіринт Морріса
Просторове навчання та пам'ять досліджувалися за допомогою завдання «Водний лабіринт Морріса» [
16]. Круглий резервуар мав діаметр 136 см, висоту 60 см і глибину 30 см, заповнений водою з температурою 24±1°C. (E), південь (S) та захід (W). Прихована кругла платформа (діаметром 10 см), занурена на 1,5 см нижче за поверхню води, розташовувалася в центрі південно-східного квадранта. Різні місця давали візуальні підказки щодо лабіринту. Затримка при втечі та пройдена відстань були виміряні для системи стеження. Щури OLETF і LETO проводили чотири випробування на день протягом п'яти днів поспіль на платформі, що знаходиться в тому самому положенні. Ми випадково призначали початкові точки щодня. Ми припиняли випробування або тоді, коли щури OLETF і щури LETO забиралися на платформу, або через 60 с. Після підйому на платформу щурам OLETF та LETO дозволяли залишатися на платформі протягом 15 с. Коли щур не знаходила платформу протягом 60 с, щур переміщали на платформу і надавали оцінку 60 с. Після завершення чотирьох випробувань щури OLETF і LETO повернулися до своїх клітин із теплим середовищем. Затримки евакуації та пройдена відстань на останній платформі були розраховані за допомогою комп'ютеризованої системи відеоспостереження (Noldus Ethovision Version 5, Wageningen, Нідерланди). Ми визначили щоденні бали як середні бали чотирьох випробувань, виконаних на день. Щури OLETF і LETO виконали 60-секундний зондовий слід із платформою, віддаленою з резервуару за даними. Час, проведений у цільовому квадранті, розраховувався з використанням комп'ютеризованої системи відеоспостереження (Noldus Ethovision Version 5, Wageningen, Нідерланди).
2.4. Біохімія крові
Коротко, експериментальних щурів умертвляли ізофлураном і збирали зразки цільної крові натще протягом ночі. Кров експериментальних щурів отримували за допомогою лівого бічного доступу калібру 20 пункції серця. Сироватка була отримана із цільної крові 9 мл у порожніх пробірках і дозволила крові повністю згорнутися. Ми центрифугували кожну пробірку у 2500 × g в протягом 15 хв і сироватку переносили в порожню пробірку для біохімічного аналізу. Ми проаналізували рівні глюкози натще за допомогою глюкометра One Touch II (Life Scan, Johnson & Johnson, Нью-Брансуік, Нью-Джерсі, США). Рівні інсуліну в сироватці, загального холестерину та тригліцеридів (TG) у сироватці були отримані за допомогою імуноферментного аналізу (ELISA; ALPCO, Салем, штат Нью-Хемпшир, США) та ферментативних наборів (Wako Chemicals USA, Inc., Річмонд, Вірджинія ). .
2.5. Аналіз клітинного циклу
Кишки щурів отримували після умертвіння, коли щурів годували протягом 8 тижнів. Клітини кишкового епітелію (IEC) виділяли з використанням Dispase (Sigma-Aldrich, Сент-Луїс, Міссурі, США) та сітчастого фільтра для клітин 45 мкм, двічі промивали PBS, фіксували 70% етанолом і фарбували йодидом пропідію (W; , USA) для аналізу клітинного сортування флуоресценції (FACS).
2.6. Фарбування гематоксиліном та еозином (H&E)
Печінка, скелетні м'язи, тонкий кишечник і шкірні тканини щурів фіксували в параформальдегіді 4% і укладали в парафінові блоки. Блоки робили на зрізи розміром 3 мкм, і зрізи фарбували гематоксиліном та еозином (H&E). Морфологічний аналіз проводився за допомогою системи аналізатора зображень Leica Qwin, оснащеної мікроскопом Leica DMLS.
2.7. Екстракція РНК та синтез кДНК
Реагент TRI від Bio-Fact (Теджон, Корея) використовували для виділення загальної РНК із 30 мг цільної тканини кишечника відповідно до інструкцій виробника. РНК кількісно визначали за допомогою Nano Drop (Nano Drop ONE, Thermo Scientific Inc., Уолтем, Массачусетс, США). Для синтезу кДНК використовували RT-Kit (M-MLV, RNase H-, Bio-Fact Co., Daejeon, Korea).
2.8. Кількісний КПЦР у реальному часі
Рівні експресії мРНК визначали за допомогою ПЛР у реальному часі (Agilent Co., Санта-Клара, Каліфорнія, США) з використанням 2X Master Mix для ПЛР у реальному часі (включаючи SYBR Green I, Low ROX, Bio-Fact Co.). Аналіз даних виконувався за допомогою програми Agilent Aria MX 1.0. Послідовності праймерів для щуриного IL6, TNFα, IL10, BiP та β-актину були наступними; (IL6) fw: TCC TAC CCC AAC TTC CAA TGC TC, rv: TTG GAT GGT CTT GGT CCT TAG CC, (TNFα) IL10) fw: ATT GAA CCA CCC GGC ATC TA, rv: AGA AAC TCC GGC GTG AGG TAG A, rv: TTT CTG GAC AGG TTT CAT GGT AG, (β - актин) fw: GGC ACC ACA CTT TCT ACA AT, rv: AGG TCT CAA ACA TGA TCT GG
2.9. Зворотня транскрипція-ПЛР
Зворотна транскрипція (RT)-PCR виконувалася з використанням 2X Taq Basic PCR Master Mix відповідно до інструкцій виробника (BioFACT Co., Корея). Зразки завантажували в агарозний гель 3% для електрофорезу і візуалізували в УФ-світлі (AE-9000 E-Graph, ATTO, Tokyo, Japan). Послідовності праймерів щуриного CHOP та гіпоксантинфосфорибозилтрансферази (HPRT) були наступними; (CHOP) fw: GTC TCT GCC TTT CGC CTT TG, rv: AGC TGG AAG CCT GGT ATG AGG A, (HPRT) fw: GCT GAC CTG CTG GAT TAC, rv: CCC GTT GAC
2.10. Статистичний аналіз
Статистичний аналіз був виконаний з використанням однофакторного дисперсійного аналізу для всіх даних та з використанням двостороннього дисперсійного аналізу з повторними вимірюваннями з наступним апостеріорним тестом Тьюкі для поведінкових даних. Результати представлені у вигляді середніх значень ± стандартні помилки, а статистична значимість була затверджена вище 0,05 від р -значення
3. Результати
3.1. Вплив вправ та добавок куркуміну на метаболічні ускладнення у щурів OLETF
Ми використовували щурів Otsuka Long-Evans Tokushima Fatty (OLETF), яка є добре відомою моделлю цукрового діабету 2 типу (T2DM), і щурів Long-Evans Tokushima Otsuka (LETO) як контроль без діабету. У щурів OLETF, отриманих з лабораторії Charles River та Дослідницького інституту Tokushima, спонтанно розвивається діабет з фенотипами поліурії, полідипсії та легкого ожиріння.
17 ]. Спочатку ми досліджували метаболізм глюкози та ліпідів у щурів OLEFT та LETO, а потім досліджували, чи вправи та добавки куркуміну позитивно впливають на щурів OLETF.
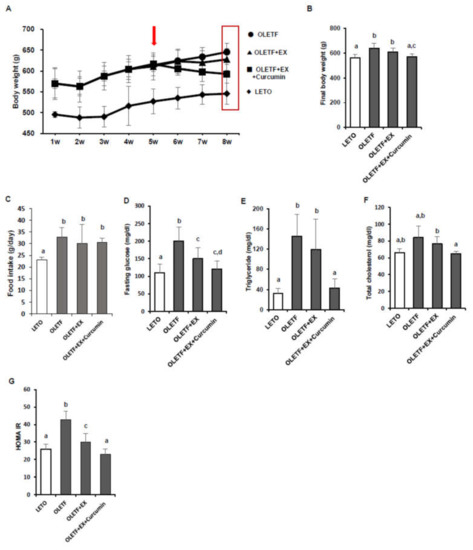
Ми виміряли зміни маси тіла, рівня глюкози натще та показників HOMA-IR ( A, B). Середня кінцева маса тіла була значно вищою для контрольних груп OLETF, ніж для контролів LETO ( p <0,05). Середня маса тіла була значно нижчою у групі OLETF + EX + куркумін, ніж у контрольній групі OLETF ( p <0,05), тоді як між контрольною групою OLETF та щурів у групі OLETF + EX не спостерігалося значної різниці в середній масі тіла ( А, Б). Середнє споживання їжі (г/день) було значно вищим у контролі OLETF, ніж у контролі LETO ( p <0,05), як і очікувалося, і не було значних відмінностей між контролю OLETF, OLETF + EX і групою OLETF + EX + куркумін ( C). Контролі OLETF мали значно вищі рівні глюкози в крові натще і HOMA-IR, ніж контролі LETO, а рівні глюкози в крові натще і HOMA-IR були значно нижчими у групах OLETF + EX та OLETF + EX + куркумін, ніж у контрольних групах OLETF і були нижче. у групі OLETF + EX + куркумін, ніж у групі OLETF + EX ( p <0,05; D, E).
Рис. 1. Фізичні вправи плюс добавка куркуміну зменшили масу тіла та покращили гомеостаз глюкози та ліпідний профіль у щурів OLETF. ( A ) Графік маси тіла (г) протягом восьми тижнів експерименту для контролю OLETF та LETO (10 самців щурів/група). Стрілка вказує на початковий період дослідження інтервенційних вправ із додаванням куркуміну. ( B ) Середня маса тіла була значно нижчою у групі OLETF + EX + куркумін, ніж у контрольній групі OLETF, і нижче у групі OLETF + EX + куркумін, ніж у групі OLETF + EX. ( C ) Споживання їжі (г/день) вимірювали двічі на тиждень. ( DРівні глюкози натще були нижче у групах OLETF + EX + куркумін та OLETF + EX, ніж у контрольній групі OLETF, але між цими групами втручання не спостерігалося значної різниці. ( E — G ) Тригліцериди сироватки, загальний холестерин та гомеостатична модель оцінки резистентності до інсуліну (HOMA-IR) у контролі OLETF та LETO. Результати представлені середні значення ± стандартне відхилення (SD). Значні відмінності визначали за допомогою однофакторного дисперсійного аналізу. Різні верхні індекси свідчить про значну різницю; а Б В Г ( р <0,05). LETO: відповідна віку контрольна група, OLETF: діабетична група, OLETF + EX: діабетична група, тренована фізичними вправами (для підйому сходами з додатковими вагами), OLETF + EX + Куркумін: діабетична група, що тренується фізичними вправами / кг) / дієта) група.
Щоб визначити, чи вправи плюс добавка куркуміну позитивний вплив на метаболізм ліпідів у щурів OLETF, були проаналізовані рівні тригліцеридів у сироватці та загального холестерину. Контролі OLETF мали значно вищі рівні тригліцеридів ( p <0,05) та загального холестерину ( p <0,05), ніж контролі LETO. Цікаво, що тригліцериди та загальний холестерин були значно нижчими у групі OLETF + EX + куркумін, ніж у контрольній групі OLETF, але не спостерігалося значної різниці між групою OLETF + EX та контролем OLETF ( F, G).
3.2. Вплив вправ та куркуміну на гістопатологію у щурів OLETF
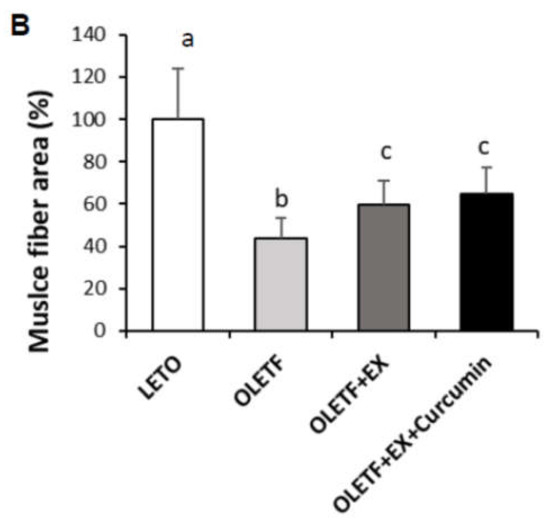
Ми вивчили вплив вправ та добавок куркуміну на морфологічні зміни у тканинах печінки та м'язів за допомогою фарбування H&E ( ). Печінкові тканини контрольних тварин OLETF показали помірну вакуолізацію, тоді як у контрольних групах LETO цього не спостерігалося. Цікаво, що у групі OLETF + EX + куркумін печінкова та м'язова тканини виглядали нормальними без вакуолізації ( A). Ні LETO, ні OLETF контролі не виявляли аномальних характеристик, таких як м'язовий фіброз литкових м'язів. Однак розміри м'язових волокон були значно меншими в контролі OLETF порівняно з контролем LETO ( p<0,05), а розмір м'язових волокон як у групі OLETF + EX, так і в групі OLETF + EX + куркумін був значно більшим у контрольній групі OLETF. У зрізах шкіри контрольні групи OLETF мали менш розвинені шари м'язів шкіри під гіподермою, ніж контролі LETO. Цікаво, що ці шари м'язів шкіри були набагато більш розвинені в групах OLETF + EX і OLETF + EX + куркумін, ніж у контрольній групі OLETF.
Рисунок 2. Куркумін знижує пов'язані з діабетом фенотипи печінки, м'язів, шкіри та тонкого кишечника у діабетичних щурів. ( A ) Типове фарбування гематоксиліном та еозином (H&E) печінки, м'язів, шкіри та тканин тонкого кишечника контрольних груп OLETF та LETO, отриманих наприкінці восьмитижневого експериментального періоду. Фізичні вправи плюс куркумін змінили морфологію печінки, м'язових шарів шкіри (вказано стрілкою) та ворсинок тонкої кишки (показано стрілкою). ( B) Кількісні дані розміру м'язових волокон експериментальних щурів. Площа поперечного перерізу м'язового волокна вимірювали всім м'язових волокон п'яти зображень, вибраних кожної тварини. У даних була розрахована середня площа м'язових волокон і представлена як середнє значення площі всіх м'язових волокон на п'яти зображеннях. Істотних відмінностей було визначено за допомогою однофакторного дисперсійного аналізу з наступним апостеріорним тестом Туреччини. Різні верхні індекси вказують на значну різницю ( p <0,05). Контроль LETO: контрольна група відповідного віку, контрольна група OLETF: діабетична група, OLETF+EX: діабетична група тренувань, OLETF+EX+ куркумін: тренування діабетиків із вправами плюс куркумін (5 г/кг у раціоні).
Вважається, що ЦД2 супроводжується хронічним запаленням, і навіть легке, але хронічне запалення може порушити епітелій тонкої кишки та коротити ворсинки. Тонкий кишечник контрольної групи OLETF показав відносно короткі та пошкоджені ворсинки порівняно з контрольною групою LETO, що дозволяє припустити, що щури OLETF мають хронічне запалення в тонкій кишці. Отже, це спостереження показало. Крім того, інфільтруючі моноцити спостерігалися у ворсинах тонкої кишки, як показано стрілками, що зазвичай спостерігається при запальних станах, таких як коліт. На фотографії, збільшеній у 40 разів, показані неправильно вирівняні інфільтровані моноцити у ворсинки групи OLETF. Цікаво, що ці явища не спостерігалися у групі OLETF+EX+ куркумін (див. A, як показано стрілками), тоді як група OLETF + EX показала менш інтенсивну інфільтрацію ворсинок моноцитами порівняно з групою OLETF, хоча ворсинки були нормальною довжиною.
3.3. Вплив вправ та куркуміну на стрес ER у тканинах тонкого кишечника щурів OLETF
Оскільки кишечник є першим бар'єром від сторонньої речовини в організмі та місцем, де куркумін всмоктується, ми вирішили досліджувати тонкий кишечник, щоб визначити, чи впливає куркумін на перше місце в організмі щодо викликаного діабетом запалення та стресу ER. Ми досліджували запальні цитокіни та рівні маркерів стресу ER у тонкому кишечнику по регіонах. Як показано на A, дванадцятипала кишка контрольних OLETF показала підвищуючу регуляцію прозапальних цитокінів (IL6 і TNFα) і знижувальну регуляцію протизапального IL10. Дивно, але вправи власними силами не усували ці ефекти, але вправи плюс куркумін значно знижували ці діабетичні ефекти, припускаючи, що вправи плюс куркумін знижували прозапальну реакцію, пов'язану з діабетом, і посилювали протизапальну реакцію. Крім того, експресія маркера стресу ER BiP ( 3А) та маркера індукованого стресом апоптозу, CHOP ( 3С) була значно нижчою у групі OLETF + EX + куркумін, ніж у контрольній групі OLETF та групі OLETF + EX. Група.
Рисунок 3. Добавка куркуміну з вправами пригнічувала прозапальні цитокіни та стрес ER у тонкому кишечнику. ( A — C ) Збирали всю дванадцятипалу кишку ( A , C ) і худу кишку ( B ), екстрагували РНК та синтезували кДНК для ОТ-ПЛР, націленої на маркер апоптозу C / EBP гомологічний білок (CHOP) ( C ) та ПЛР в реальному часі, націлена на прозапальні цитокіни (IL6 та TNFα (прозапальні цитокіни), IL10 (протизапальний цитокін)) та маркер стресу ER BiP ( A , B ). Гіпоксантинфосфорибозилтрансфераза (HPRT) і β-актин використовувалися як контроль завантаження для ОТ-ПЛР та ПЛР в реальному часі відповідно. (D ) У контрольній групі OLETF куркумін у поєднанні з вправами рятували фазу G0/G1 клітинного циклу в ізольованих клітинах тонкого кишечника. Ізольовані клітини тонкої кишки фіксували етанолом 70% і фарбували йодидом пропідію (PI). Проточну цитометрію використовували для кількісного визначення клітинних фаз (% від осі Y). Істотних відмінностей було визначено з використанням однофакторного дисперсійного аналізу з наступним апостеріорним тестом Туреччини. Різні верхні індекси вказують на значну різницю ( p <0,05). Результати представлені як середніх значень ± стандартні помилки. Контрольна група LETO: відповідна віку контрольна група, контрольна група OLETF: діабетична група, група OLETF + EX: група діабетиків, тренована фізичними вправами, OLETF + EX + куркумін: група діабетиків, тренована фізичними вправами, яка одержувала куркумін ).
Аналогічна картина спостерігалася і в худій кишці. Як показано на 3B рівні IL6 і TNFα були значно вищими в контролі OLETF, ніж у контролі LETO, але цей ефект значно зменшувався при використанні вправ плюс куркумін більшою мірою, ніж при використанні тільки вправ. Крім того, BiP був значно нижчим у групі OLETF + EX + куркумін, ніж у групі OLETF + EX та у контрольній групі OLETF. Ці дані показують, що вправи у поєднанні з додаванням куркуміну вплинули на запалення, пов'язане з діабетом, і стрес ER, але ці вправи самі по собі не мали такого помітного ефекту.
Крім того, ми досліджували, чи відрізняється клітинний цикл клітин тонкої кишки у контролі OLETF та LETO. Ми спостерігали, що фаза G0/G1 клітинного циклу була ослаблена в групі OLETF + EX та контролі OLETF у порівнянні з контролем LETO ( D), але що фаза G0 / G1 була вищою в групі OLETF + EX + куркумін, ніж у групі OLETF + EX і в контролі OLETF ( p <0,05). Ці спостереження припускають, що знижені рівні мРНК CHOP та рівні мРНК BiP, викликані вправами плюс додавання куркуміну, можуть збільшувати відсоток клітинного циклу G0/G1 на стадії виживання або проліферації клітин та, таким чином, покращувати когнітивні функції у щурів OLETF.
3.4. Вплив вправ та добавок куркуміну на навчання та дефіцит пам'яті у щурів OLETF
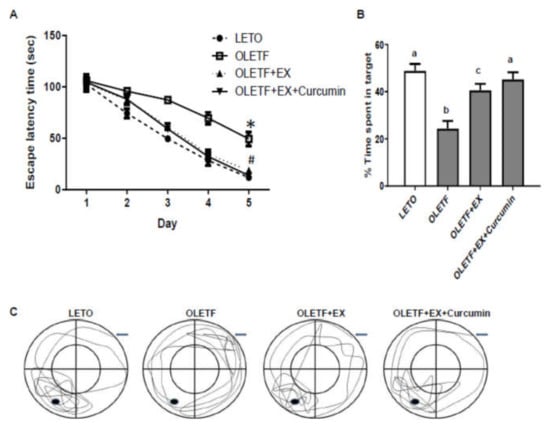
Когнітивні функції оцінювалися за допомогою тесту водного лабіринту Морріса. Середня латентність втечі зменшилася протягом послідовних п'яти днів періоду навчання у всіх групах ( 3А). Ми виявили, що середня латентність втечі була більше для OLETF, ніж для контролю LETO (49,3 проти 11,8 с у щурів OLETF в порівнянні з щурів LETO в системі стеження, p <0,05), але значно менше групи OLETF + EX + куркумін (13,8 з). ніж у групі OLETF + EX (18,8 с) та у контролі OLETF (49,4 с) ( A).
Рисунок 4. Куркумін у поєднанні з вправами покращив навчання та збереження пам'яті у щурів OLETF у тесті у водному лабіринті Морріса. ( A ) Затримка втечі вимірювалася із встановленою платформою. По чотири випробування на день експериментальні щури виконували для отримання даних протягом п'яти днів поспіль. * p <0,05 порівняно з контрольними пацюками LETO, # p <0,05 порівняно з пацюками OLETF у контролі. ( B — C ) Вплив вправ та куркуміну на збереження пам'яті у випробуванні з зондом. Час, проведений у цільовому квадранті у випробуванні зонда, розрахували на шостий день ( B ). Типова діаграма трекінгу по довжині плавальної доріжки. Дані були визначені за допомогою системи відеоспостереження ( CКонтрольна група LETO: відповідна віку контрольна група, контрольна група OLETF: діабетична група, OLETF+EX група: діабетична група тренувань, OLETF+EX+ куркумін група: діабетична група тренувань та куркуміну (дієта 5 г/кг). Результати представлені як середніх значень ± стандартні помилки. Статистичний аналіз проводився з використанням однофакторного дисперсійного аналізу для пробного дослідження та з використанням двостороннього дисперсійного аналізу з повторними вимірами з наступним апостеріорним тестом Туреччини для поведінкових даних. Різні верхні індекси вказують на значну різницю ( p <0,05).
Для оцінки збереження пам'яті ми провели випробування зонда через два дні після останнього тесту у водному лабіринті Морріса. Пацюків допускали до плавання протягом 60 с у басейні, в якому вони тренувалися, але з прибраною платформою для порятунку. Відсоток часу плавання, проведеного в межах цільового квадранта у контрольних груп OLETF, був меншим, ніж у контрольних груп LETO (24,4% проти 48,9% щурів OLETF порівняно з щурами LETO у системі стеження, p <0,05) і щурами в OLETF+. EX (40,6%) і OLETF + EX + куркумін (45,2%) і був значно меншим у групі OLETF + EX, ніж у групі OLETF + EX + куркумін ( B, C). Ці результати показують, що вправи плюс куркумін можуть покращити когнітивні функції.
4. Обговорення
У цьому дослідженні ми припустили, що комбінація вправ та куркуміну може синергетично послаблювати запальну реакцію при діабеті. Наші результати показують, що вправи разом із добавкою куркуміну послаблювали діабетичні фенотипи у щурів OLETF, що визначалося вимірами маси тіла, глюкози, інсуліну, тригліцеридів та загального холестерину. Зокрема, ми виявили, що вправи з куркуміном підвищують чутливість до інсуліну, як визначено індексами HOMA-IR, та знижують рівень глюкози натще у щурів OLETF.
Запалення - це фундаментальний біологічний процес, який відновлює гомеостаз тканин за допомогою різних механізмів відновлення та впливає на багато гострих та хронічних патологічних станів. Однак правильне регулювання запальних механізмів важливе для запобігання неконтрольованому посиленню початкових запальних реакцій та переходу від відновлення тканин до побічних ушкоджень та розвитку захворювань [
18 ]. Отже, легке хронічне запалення є найпоширенішим фенотипом метаболічного синдрому, включаючи ЦД2. Було показано, що стрес ER відіграє важливу роль, а також є звичайним явищем між запаленням та ЦД2, а також багатьма іншими метаболічними порушеннями [
19]. Хоча причини, що призводять до стресу ER, досі не визначені, передбачається, що причиною є підвищення циркулюючих прозапальних цитокінів, таких як TNFα та IL6, у цих метаболічних умовах.
8 ]. При T2DM посилення передачі сигналів IRE1α активує шлях NF-kB шляхом фосфорилювання N-кінцевої кінази 1 c-jun (JNK), добре відомих модуляторів запалення, що сприяють експресії генів, і активності білків, що беруть участь у регуляції запальної реакції. Це також призводить до порушення передачі сигналів інсуліну та активації декількох молекул у скелетних м'язах, жировій тканині та печінці.
8 ]. Повідомлялося, що порушення передачі сигналів інсуліну в результаті стресу в ER призводить до системної інсулінорезистентності, гіперглікемії та гіперінсулінемії.
7]. У дослідженні наші результати з використанням щурів OLETF, природно індукованої моделі СД2, підтвердили підвищену інсулінорезистентність, представлену як HOMA-IR на контрольній групі OLETF, і що цей фенотип був покращений за допомогою комбінованої терапії вправ та добавок куркуміну.
Діабет є прикладом патологічного нейродегенеративного процесу, пов'язаного із хронічними запальними змінами. Відомо, що основні механізми діабету та когнітивного дефіциту включають підвищені рівні фосфорильованого тау-білка (p-Tau) у спинномозковій рідині та тривалий некодуючий РНК-індукований апоптоз нейронів гіпокампу у літніх діабетичних тварин та пацієнтів [
20 ,
21 ]. Декілька досліджень показали, що вправи зменшують запальну реакцію мозку при діабеті [
22 ,
23 ,
24 ,
25 ,
26]. Однак інші дослідження на людях показали, що зміни одного фактора способу життя, такі як зміни фізичної активності, меншою мірою впливають на когнітивні функції при нейродегенеративних захворюваннях. Куркумін є добре відомим природним антиоксидантом, який знижує запальну реакцію при діабеті через сигнальні шляхи Nrf2 та PI3K/AKT [
27 ,
28 ,
29 ,
30 ,
31 ]. Дані показують, що куркумін може покращувати патології, пов'язані з амілоїдом-β, і відновлювати патології, пов'язані з p-тау-білком, на тваринних моделях [
32 ,
33 ]. Також було показано, що куркумін знижує експресію гена рецептора інсуліну та субстрату-1 рецептора інсуліну в гіпокампі [
34 ,
35 ,
36 ]. Відповідно, наші результати показують, що комбінація вправ та куркуміну синергетично покращує когнітивні функції у щурів OLETF або запальну реакцію у кишечнику OLETF.
У моделі мишей зі зниженим стресом ER, специфічної для мозку, важка резистентність до лептину значно збільшувала ожиріння на дієті з високим вмістом жирів, що передбачає, що стрес ER може брати участь у регулюванні чутливості до інсуліну в мозку [
37 ]. У багатьох попередніх дослідженнях ми припустили, що стрес ER, який спостерігається в кишечнику в нашому дослідженні, також відбувається в мозку, оскільки стрес ER виникає в різних тканинах системно, і постулювали, що пригнічення стресу ER кишечника за допомогою вправ та добавок куркуміну відбудеться у тканину головного мозку, припускаючи, що непряме придушення стресу ER у головному мозку може покращити когнітивні функції мозку.
18 ,
19 ,
38 ,
39 ,
40 ,
41].
Нові доклінічні дані показали, що двонаправлена передача сигналів між шлунково-кишковим трактом (ЖКТ) та мозком, так звана вісь кишечник-мозок, відіграє важливу роль як у метаболізмі, так і у поведінці господаря [
42 ]. Наявні дані свідчать про те, що мікробна різноманітність знижується при порушенні регуляції метаболізму.
43 ,
44 ,
45 ]. Виявлено, що мікробні метаболіти коротколанцюгових жирних кислот мають безліч фізіологічних ефектів, включаючи енергетичний гомеостаз, за рахунок регуляції гормонів ШКТ, таких як холецистокінін, глюкагоноподібний пептид 1 (GLP1) та лептин [
46]. Більш того, кишкові мікробіоми викликають зміни в секреції кишкових гормонів та імунній відповіді, а також у бар'єрі слизової оболонки кишечника, контролі гематоенцефалічного бар'єру та створенні та розкладанні нейромедіатора, таким чином, тісно взаємодіючи з центральною нервовою системою, що впливає на здатність мозку розвивати. . Кореляції між мікробіомом кишечника та так званою віссю мозок-кишковик інтенсивно вивчаються при неврологічних захворюваннях. Одне дослідження показало, що втрата толл-подібного рецептора (TLR) 2, одного з датчиків вродженого імунітету, у традиційних мишей призводить до фенотипу метаболічного синдрому, що нагадує, що характеризується чіткою відмінністю мікробіоти кишечника, що викликає резистентність до інсуліну. та запалення, пов'язане зі стресом ER,
47 ]. Ці дані підкреслили роль мікробіоти у складній мережі молекулярних та клітинних взаємодій, які пов'язують генотип з фенотипом та мають потенційні наслідки для широкого спектру поширених захворювань людини, включаючи діабет, та навіть інших імунологічних порушень.
43 ].
Проте це дослідження було кілька обмежень. По-перше, дослідження було зосереджено на вплив вправ щурів OLETF на когнітивну функцію, яка не забезпечує експериментальній групі щурів OLETF лише куркумін. Тим не менш, вправи є основним фактором способу життя дорослих щодо когнітивної функції, що також було включено до дослідження для вивчення впливу куркуміну. По-друге, ми не надали дані гістології чи ER стресу у головному мозку. Однак наша лабораторія раніше повідомляла, що у щурів OLEF з вправами в гіпоталамусі спостерігалося значне підвищення рівнів мРНК GLP-1 (основного інкретинового гормону), переносника глюкози 2 (GLUT2) та супероксиддисмутази 1 (SOD1, антиоксидантний фермент) у порівнянні з Щура OLE мозку [
48]. Справді, як згадувалося вище, ми виявили, що вправи у поєднанні з куркуміном усувають стресову реакцію ER у тканинах кишечника контрольної групи OLETF та підвищують чутливість до інсуліну у системі крові. Крім того, щури в групі OLETF + EX + куркумін продемонстрували кращі навички навчання та пам'ять мозку у тесті у водному лабіринті Морріса, який передбачає, що вправи плюс куркумін регулюють чутливість до інсуліну у всьому організмі, знижуючи стрес ER та підвищуючи резистентність до . В результаті рівень інсуліну регулюється, стрес ER, спричинений високим рівнем глюкози в крові, знижується, а апоптоз, спричинений стресом ER, пригнічується в системі крові, а когнітивні функції покращуються у щурів OLETF. Необхідні подальші дослідження для підтвердження результатів у різних тканинах, включаючи мозок, з точки зору запалення та стресу ER, щоб зменшити симптоми діабетичної хвороби.
Як третє обмеження куркумін, як біоактивне з'єднання, розчинний в етанолі, лугу, оцтовій кислоті та інших органічних розчинниках і погано всмоктується в кишечнику. В одному дослідженні повідомлялося про всмоктування 400 мг куркуміну, введеного перорально щурам, і було виявлено у 5 мг/л у ворітній вені печінки через 24 години після пероральної ін'єкції.
49 ,
50 ].
Однак у цьому дослідженні ми спостерігали, що щури, які отримували куркумін у кількості 5 г на раціон (кг), покращували втрату ваги, гомеостаз глюкози та ліпідний профіль. Незважаючи на ці обмеження, наявні дані дозволяють зробити висновок, що поєднання вправ і куркуміну покращує когнітивні явища, пов'язані з діабетом, у щурів.
Внесок авторів
Концептуалізація, EP, JAC та JHY; методологія, SHP; ресурси, EP та JAC; курування даних, JC та J.-OK; Підготовка оригінального чернетки, JAC та EP; фінансування залучення, EP та JAC Усі автори прочитали та погодилися з опублікованою версією рукопису.
Фінансування
Це дослідження було підтримане Програмою фундаментальних наукових досліджень через Національний дослідний фонд Кореї, що фінансується Міністерством освіти: NRF-2019R1H1A2039746 (EP) та NRF-2019R1A2B5B01070133 (JAC).
Подяки
Ми дякуємо Хену Лі, Хелім Пак, Хе-Чон Хван і Джу-Ген Юн за їхню допомогу у всіх експериментах на тваринах.
Конфлікт інтересів
Автори не заявляють про конфлікт інтересів.
Посилання
- Біссельс, ГДж; Реймер, Ю. Д. Зміни мозку, що лежать в основі когнітивної дисфункції діабету: чому ми можемо навчитися за допомогою МРТ? Діабет 2014 , 63 , 2244-2252. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Біссельс, ГДж; Страчан, MWJ; Visseren, FL; Каппелле, LJ; А. Вітмер, Р. Деменція та зниження когнітивних функцій при діабеті 2 типу та передіабетичних стадіях: на шляху до цільових заходів. Ланцет Діабет Ендокринол. 2014 , 2 , 246-255. [ Google Scholar ] [ CrossRef ]
- Riederer, P.; Корчин, А.Д.; Алі, СС; Bajenaru, O.; Choi, MS; Чопп, М.; Дерманович-Доброта, В.; Grünblatt, E.; Jellinger, KA; Камаль, Массачусетс; та інші. Діабетичний мозок та пізнання. J. Neural Transm. 2017 , 124 , 1431-1454. [ Google Scholar ] [ CrossRef ]
- Чжао, Р.Р.; О'Салліван, AJ; Сінгх, MAF. Вправи або фізична активність та когнітивні функції у дорослих з діабетом 2 типу, інсулінорезистентністю або порушеною толерантністю до глюкози: систематичний огляд. Євро. Rev. Aging Phys. Закон. 2018 , 15 , 1. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Хардінг, HP; Zhang, Y.; Ron, D. Трансляція та фолдинг білка пов'язані з кіназою, резидентною в ендоплазматичному ретикулумі. Nature 1999 , 397 , 271-274. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hwang, J.; Ци, Л. Контроль якості в ендоплазматичній мережі: перехресні перешкоди між шляхами ERAD та UPR. Trends Biochem. Sci. 2018 , 43 , 593-605. [ Google Scholar ] [ CrossRef ]
- Ozcan, U.; Yilmaz, E.; Ozcan, L.; Furuhashi, M.; Vaillancourt, E.; Сміт, РВ; Гергюн, Чехія; Хотамислигіл, Г.С. Хімічні шаперони зменшують стрес ER та відновлюють гомеостаз глюкози на мишачій моделі діабету 2 типу. Science 2006 , 313 , 1137-1140. [ Google Scholar ] [ CrossRef ]
- Хотамислигіл, Г.С. Стрес ендоплазматичного ретикулуму та запальна основа метаболічних захворювань. Cell 2010 , 140 , 900-917. [ Google Scholar ] [ CrossRef ]
- Chen, R.; Shi, J.; Інь, Q.; Li, X.; Sheng, Y.; Han, J.; Zhuang, P.; Чжан Ю. Морфологічні та патологічні характеристики мозку при діабетичній енцефалопатії. J. Alzheimers Dis. 2018 , 65 , 15-28. [ Google Scholar ] [ CrossRef ]
- Чжан, X.; Xu, L.; Він д .; Лінг, С. Ендоплазматичний ретикулум, спричинений стресом, апоптоз нейронів гіпокампу, залучений до діабетичного когнітивного порушення. BioMed Res. Int. 2013 , 2013 , 1-9. [ Google Scholar ] [ CrossRef ]
- Чо, JA; Park, E. Куркумін використовує шлях протизапальної реакції для захисту кишківника від бактеріальної інвазії. Nutr. Res. Практик. 2015 , 9 , 117-122. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Чой, Ю.Є.; Park, E. Куркумін підвищує чутливість інгібітору полі (АДФ-рибози) полімерази до хіміотерапії у клітинах раку грудей. J. Nutr. Biochem. 2015 , 26 , 1442-1447. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Бугліані, М.; Mossuto, S.; Grano, F.; Сулейман, М.; Marselli, L.; Boggi, U.; De Simone, P.; Ейзірік, DL; Cnop, M.; Marchetti, P.; та інші. Модуляція аутофагії впливає на функцію та виживання бета-клітин підшлункової залози людини в умовах стресу ендоплазматичної мережі та при діабеті 2 типу. Фронт. Ендокринол. 2019 , 10 , 52. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kawano, K.; Hirashima, T.; Mori, S.; Saitoh, Y.; Куросумі, М.; Нотори, Т. Спонтанна тривала гіперглікемія у щурів з діабетичними ускладненнями: штам Оцука Лонг-Еванс Токусіма жирний (OLETF). Діабет 1992 , 41 , 1422-1428. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kim, H.-J.; Сонг, У. Тренування з обтяженнями збільшує рівень фактора зростання фібробластів-21 та рівень ірізину в скелетних м'язах щурів з діабетом Цукера з ожирінням. J. Exerc. Nutr. Biochem. 2017 , 21 , 50-54. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Vorhees, CV; Вільямс, М. Т. Водний лабіринт Морріса: процедури для оцінки просторових та пов'язаних форм навчання та пам'яті. Nat. Protoc. 2006 , 1 , 848-858. [ Google Scholar ] [ CrossRef ]
- Kawano, K.; Hirashima, T.; Mori, S.; Нотори, Т. OLETF (Otsuka Long-Evans Tokushima Fatty) щур: нова лінія щурів з NIDDM. Diabetes Res. Clin. Практик. 1994 , 24 , S317 - S320. [ Google Scholar ] [ CrossRef ]
- Sims-Robinson, C.; Zhao, S.; Hur, J.; Фельдман, Е. Л. Стрес ендоплазматичного ретикулуму центральної нервової системи на мишачій моделі діабету 2 типу. Діабетол. 2012 , 55 , 2276-2284. [ Google Scholar ] [ CrossRef ]
- Чжан, X.; Zhang, G.; Zhang, H.; Карін, М.; Bai, H.; Цай, Д. Гіпоталамічний IKKβ/NF-κB та ER стрес пов'язують переїдання з енергетичним дисбалансом та ожирінням. Cell 2008 , 135 , 61-73. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Yu, J.; Li, C.; Че, Л.; Zhao, Y.; Guo, Y. Пригнічення довгою некодуючою РНК H19 рятує нейрони гіпокампа від апоптозу та окисного стресу, інгібуючи метилювання IGF2 у мишей з цукровим діабетом, індукованим стрептозотоцином. J. Cell. Physiol. 2018 , 234 , 10655-10670. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Moran, C.; Beare, R.; Phan, TG; Брюс, Д.Г.; Каллісая, М.; Срікант, В.К. Цукровий діабет 2 типу та біомаркери нейродегенерації. Неврологія 2015 , 85 , 1123-1130. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Белотто, МФ; Magdalon, J.; Родрігес, HG; Виноло, МАР; Curi, R.; Піфон-Курі, ТЗ; Хатанака, Еге. Помірні вправи покращують функцію лейкоцитів і зменшують запалення при діабеті. Clin. Exp. Імунол. 2010 , 162 , 237-243. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Karstoft, K.; Педерсен, Б.К. Вправи та діабет 2 типу: акцент на метаболізм та запалення. Імунол. Cell Boil. 2015 , 94 , 146-150. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- De Lemos, ET; Пінто, РМ; Oliveira, J.; Garrido, P.; Sereno, J.; Mascarenhas-Melo, F.; Пінейро, JP; Тейшейра, Ф.; Рейс, Ф. Диференціальні ефекти гострих (послаблюючих) та хронічних (тренувальних) вправ на запалення та статус окисного стресу на тваринній моделі цукрового діабету 2 типу. Mediat. Запалення. 2011 , 2011 , 1-8. [ Google Scholar ] [ CrossRef ]
- Waddington, G. Інтенсивність вправ та запалення при діабеті 2 типу. J. Sci. Med. Спорт 2017 , 20 , 885. [ Google Scholar ] [ CrossRef ]
- Zoppini, G.; Targher, G.; Zamboni, C.; Вентурі, C.; Cacciatori, V.; Moghetti, P.; Muggeo, M. Вплив тренувань середньої інтенсивності на біомаркери запалення та ендотеліальної дисфункції у літніх пацієнтів із діабетом 2 типу. Nutr. Метаб. Кардіоваськ. Dis. 2006 , 16 , 543-549. [ Google Scholar ] [ CrossRef ]
- Biswas, S.; Chen, S.; Liang, G.; Feng, B.; Cai, L.; Хан, З.А.; Чакрабарті, С. Аналоги куркуміну знижують показники стресу та запалення в експериментальних моделях діабету. Фронт. Ендокринол. 2019 , 10 , 887. [ Google Scholar ] [ CrossRef ]
- Ghosh, S.; Banerjee, S.; Sil, PC Сприятлива роль куркуміну при запаленні, діабеті та нейродегенеративних захворюваннях: нещодавнє оновлення. Food Chem. Toxicol. 2015 , 83 , 111-124. [ Google Scholar ] [ CrossRef ]
- Рашид, К.; Chowdhury, S.; Ghosh, S.; Sil, PC Куркумін послаблює спричинене окислювальним стресом запалення, опосередковане NFκB, та залежний від ендоплазматичного ретикулуму апоптоз спленоцитів при діабеті. Biochem. Pharmacol. 2017 , 143 , 140-155. [ Google Scholar ] [ CrossRef ]
- Weisberg, S.; Leibel, R.; Торторієлло, Д.В. Дієтичний куркумін значно знижує запалення, пов'язане з ожирінням, та діабет на мишачих моделях діабету. Ендокринологія 2008 , 149 , 3549-3558. [ Google Scholar ] [ CrossRef ]
- Ян, З.; Dai, Y.; Fu, H.; Zheng, Y.; Bao, D.; Інь, Й.; Chen, Q.; Не, X.; Hao, Q.; Hou, D.-R.; та інші. Куркумін має захисну дію проти передчасної недостатності яєчників у мишей. J. Mol. Ендокринол. 2018 , 60 , 261-271. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Вітання.; Wang, P.; Wei, P.; Feng, H.; Ren, Y.; Yang, J.; Rao, Y.; Shi, J.; Тіан, Дж. Вплив куркуміну на синапси у мишей APPswe/PS1dE9. Int. J. Immunopathol. Pharmacol. 2016 , 29 , 217-225. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Ma, Q.-L.; Zuo, X.; Ян, Ф.; Переконання, О. Дж.; Гант, діджей; Алавердян, М.; Teng, E.; Hu, S.; Chen, P.-P.; Maiti, P.; та інші. Куркумін пригнічує розчинні димери тау-білка та коригує молекулярні шаперони, синаптичні та поведінкові дефіцити у старих трансгенних мишей людини. J. Boil. Chem. 2012 , 288 , 4056-4065. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Feng, H.-L.; Dang, H.-Z.; Fan, H.; Chen, X.-P.; Рао, Y.-X.; Ren, Y.; Yang, J.-D.; Shi, J.; Wang, P.; Тіан, Ж.-З. Куркумін покращує сигнальний шлях інсуліну в мозку трансгенних мишей із хворобою Альцгеймера. Int. J. Immunopathol. Pharmacol. 2016 , 29 , 734-741. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zhang, L.; Kong, X.-J.; Wang, Z.-Q.; Xu, F.-S.; Чжу, Ю.-Т. Дослідження нейропротекторної дії куркуміну на мозок щурів із діабетом. J. Nutr. Зцілити. Старіння 2016 , 20 , 835-840. [ Google Scholar ] [ CrossRef ]
- Wang, P.; ВС ц .; Feng, H.; Чен, X.; Dong, Y.; Rao, Y.; Ren, Y.; Yang, J.; Shi, J.; Tian, J.; та інші. Куркумін регулює шляхи інсуліну та метаболізм глюкози в мозку мишей APPswe/PS1dE9. Int. J. Immunopathol. Pharmacol. 2017 , 30 , 25-43. [ Google Scholar ] [ CrossRef ]
- Ozcan, L.; Ергін, АС; Лу, А.; Chung, J.; Sarkar, S.; Nie, D.; Майєрс, MG; Озкан, У. Стрес ендоплазматичного ретикулума грає центральну роль розвитку резистентності до лептину. Cell Metab. 2009 , 9 , 35-51. [ Google Scholar ] [ CrossRef ]
- Бускетс, О.; Eritja, À.; Лопес, Б.М.; Ettcheto, M.; Manzine, P.; Кастро-Торрес, РД; Verdaguer, E.; Olloquequi, J.; Vázquez-Carrera, M.; Auladell, C.; та інші. Роль N-кінцевої кінази 2 мозку c-Jun у контролі рецептора інсуліну та її зв'язок з когнітивними функціями у доклінічній моделі дієти з високим вмістом жирів. J. Neurochem. 2019 , 149 , 255-268. [ Google Scholar ] [ CrossRef ]
- Kong, F.-J.; Торговий центр.; Guo, J.-J.; Xu, L.-H.; Li, Y.; Ку, S. Шлях стресу/аутофагії ендоплазматичного ретикулуму бере участь у індукованому діабетом апоптозі нейронів та зниженні когнітивних функцій у мишей. Clin. Sci. 2018 , 132 , 111-125. [ Google Scholar ] [ CrossRef ]
- Muriach, M.; Флорес-Беллвер, М.; Romero, FJ; Барсія, Дж. М. Діабет та мозок: окисний стрес, запалення та аутофагія. Oxidative Med. Cell. Лонгів. 2014 , 2014 , 1-9. [ Google Scholar ] [ CrossRef ]
- Zhao, Y.; Ян, Ю.; Zhao, Z.; Li, S.; Інь, Дж. Динамічні зміни маркерів шляху стресу ендоплазматичного ретикулуму GRP78 та CHOP у гіпокампі мишей з діабетом. Brain Res. Бик. 2015 , 111 , 27-35. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Cryan, JF; Дінан, Т.Г. Мікроорганізми, що змінюють свідомість: вплив мікробіоти кишечника на мозок і поведінку. Nat. Rev. Neurosci. 2012 , 13 , 701-712. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Де Клерк, Північна Кароліна; Фрісен, Міннесота; Groen, AK; Nieuwdorp, M. Мікробіота кишечника та вісь кишечника та мозку. Психосом. Med. 2017 , 79 , 874-879. [ Google Scholar ] [ CrossRef ]
- Arora, T.; Бекхед, Ф. Мікробіота кишечника та метаболічні захворювання: поточне розуміння та перспективи на майбутнє. J. Intern. Med. 2016 , 280 , 339-349. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Ianiro, G.; Bibbò, S.; Антоніо, G.; Каммарота, Г. Терапевтична модуляція мікробіоти кишечника: поточні клінічні застосування та перспективи на майбутнє. Curr. Drug Targets 2014 , 15 , 762-770. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Yadav, H.; Lee, J.-H.; Lloyd, J.; Walter, P.; Рейн, С.Г. Сприятливі метаболічні ефекти пробіотика через секрецію гормону GLP-1, спричинену бутиратом. J. Boil. Chem. 2013 , 288 , 25088-25097. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Каріциллі, AM; Пікарді, ПК; Де Абре, LL; Ueno, M.; Prada, PO; Мотузка, ER; Hirabara, SM; Castoldi, A.; Де Мораєш-Вієра, PMM; Камара, н. та інші. Мікробіота кишечника є ключовим модулятором інсулінорезистентності мишей з нокаутом TLR 2. PLoS Кип'ятити. 2011 , 9 , e1001212. [ Google Scholar ] [ CrossRef ]
- Парк, Ш; Юн, JH; Seo, DY; Кім, TN; Ko, J.; Хан, Дж. Тренування з обтяженнями знижує втрату ендогенного рецептора GLP-1 у гіпоталамусі щурів із діабетом 2 типу. Int. J. Environ. Res. Public Health 2019 , 16 , 830. [ Google Scholar ] [ CrossRef ]
- Равіндранатх, V.; Чандрасекхара, Н. Поглинання та розподіл куркуміну в тканинах у щурів. Toxicol. 1980 , 16 , 259-265. [ Google Scholar ] [ CrossRef ]
- Xue, M.; Cheng, Y.; Xu, L.; Чжан, Л. Дослідження характеристик абсорбції куркуміну в кишечнику in vivo та in vitro. J. Appl. Pharm. 2017 , 9 . [ Google Scholar ] [ CrossRef ]
© 2020 Автори. Ліцензіат MDPI, Базель, Швейцарія. Ця стаття — це стаття у відкритому доступі, яка розповсюджується відповідно до умов ліцензії Creative Commons Attribution (CC BY) (http://creativecommons.org/licenses/by/4.0/ )




Без коментарів