1. Введение
Диабет — это заболевание, при котором нарушение регуляции передачи сигналов инсулина приводит к инсулинорезистентности [
1 ]. Клинические исследования показали, что гликемическая регуляция и нарушение выделения инсулина отрицательно влияют на функцию мозга [
2 ]. Кроме того, несколько исследований показали, что диабет является фактором риска когнитивных заболеваний, таких как сосудистая деменция и болезнь Альцгеймера [
3 ]. Соответственно, пожилые пациенты с диабетом 2 типа (СД2) получают клинические рекомендации о положительном влиянии упражнений на повышение чувствительности к инсулину [
4 ]. Однако несколько всеобъемлющих руководств продемонстрировали связь между когнитивной дисфункцией и диабетом.
Эндоплазматический ретикулум (ЭР) играет важную роль в синтезе белков, а также в сворачивании и процессинге секреторных и трансмембранных белков [
5 ]. Когда неправильно свернутые или развернутые белки накапливаются в ER из-за генетических факторов или факторов окружающей среды, возникает стресс ER, который опосредуется тремя трансмембранными белками ER, IRE1, PERK и ATF6, вызывая ответ развернутых белков (UPR) [
6 ].
Поскольку стресс ER может активировать клеточные воспалительные пути, которые нарушают клеточные функции и приводят к метаболическим нарушениям, существующие данные свидетельствуют о том, что стресс ER является ключевым звеном между T2DM, ожирением и инсулинорезистентностью [
7 ,
8 ]. Известно, что воздействие высоких уровней глюкозы может вызвать стресс ER за счет генерации свободных радикалов, аберрантного гликозилирования белка и увеличения мембранного и белкового обмена, что приводит к усилению экспрессии белка гомологии C / EBP (CHOP) (известный медиатор стресса ER -индуцированный апоптоз).
Диабет также связан с церебральной атрофией, неврологическими нарушениями, снижением когнитивных функций и психомоторной дисфункцией, называемой диабетической энцефалопатией (ДЭП) [
9 ]. Растущее количество данных указывает на то, что окислительный стресс, хроническое воспаление, митохондриальная дисфункция, нарушение передачи сигналов инсулина и стресс ER способствуют развитию DEP. Однако мало исследований изучали влияние кишечного стресса ER.
Сообщалось, что в начале стресса ER повышенная экспрессия GRP78 в головном мозге крысы предотвращала индуцированный CHOP апоптоз, но когда стресс ER был устойчивым, уровни CHOP заметно повышались в гиппокампе крыс с диабетом [
10 ]. Это предполагает, что апоптоз, опосредованный стрессом CHOP-ER, может быть вовлечен в индуцированные гипергликемией синапсы гиппокампа и нейрональные нарушения и, таким образом, способствовать диабетическим когнитивным нарушениям.
Куркумин — это природное соединение с известными антиоксидантными и противовоспалительными свойствами [
11 ,
12 ]. Хотя было показано, что он вызывает апоптоз раковых клеток, его влияние на воспалительную реакцию при СД2 недостаточно изучено. Накапливающиеся данные указывают на то, что слабое воспаление, окислительный стресс и стресс ER способствуют резистентности к инсулину при СД2. Следовательно, похоже, что подавление стресса и воспаления ER может улучшить когнитивные функции при СД2 [
13 ]. Однако не было установлено, могут ли упражнения в сочетании с добавлением куркумина улучшить когнитивную дисфункцию, связанную с диабетом, путем регулирования стрессовой реакции ER.
В настоящем исследовании мы исследовали, различаются ли когнитивные функции у крысы Otsuka Long-Evans Tokushima Fatty (OLETF) (крыса с диабетом) и у Long-Evans Tokushima Otsuka (LETO; крыса из контроля), и существует ли комбинация упражнения и добавка куркумина улучшат когнитивную дисфункцию, связанную с диабетом. Кроме того, мы измерили изменения в уровнях цитокинов воспалительного ответа и CHOP, вызванные комбинацией упражнений и добавок куркумина в модели спонтанного диабета на крысах.
2. Материалы и методы
2.1. Животные и экспериментальный дизайн
В качестве контроля использовали крыс LETO (контрольные самцы LETO;
n = 10), а в качестве модели СД2 использовали крыс OLETF, гиперфагический фенотип с мутациями рецептора холецистокинина-1 [
14 ]. Самцов крыс LETO и OLETF содержали по два в клетке при температуре 20 ± 2,5 ° C и 12:12 часовом цикле свет-темнота в светлом помещении, снабжали достаточным питанием и водой, а также контролировали массу тела и потребление пищи. Крыс кормили либо стандартной пищей (SC; 10 ккал% жира, диета D12492 соответствует 7% сахарозы, исследовательская диета, США; контроль OLETF), либо стандартной диетой (SC), содержащей куркумин (диета 5 г / кг, Sigma, Каталожный номер C1386). Крысы OLETF были случайным образом разделены на три группы (контрольная группа OLETF;
n = 10), диабетическая группа, выполнявшая упражнения (группа OLETF + EX;
n= 10), или группа диабетиков, которые тренировались плюс куркумин (группа OLETF + EX + куркумин;
n = 10) в возрасте 22 недель. Самцы крыс LETO были сопоставлены по возрасту. Все результаты экспериментов на животных были одобрены Комитетом по использованию и уходу за животными в Университете Ханнам (HNU2016-16).
2.2. Протокол упражнений
Крыс в группе OLETF + EX обучали подниматься по вертикальной лестнице длиной 1 м (наклон 85 °) с грузами, прикрепленными к их хвостам. В течение первой недели крыс приучали забираться на верхнюю клетку с грузом или без него. Тренировочные занятия со второй недели начинали с весом 50% от массы тела. Крысы начали подниматься с нижней части лестницы и были вынуждены подняться наверх. Когда они достигли вершины, им дали 2-х минутный отдых перед следующим испытанием. Последующие испытания проводили снизу лестницы, и при каждом испытании к предыдущему весу добавляли 20 г. Если крыса могла подняться 10 раз с увеличением веса, тренировка считалась завершенной. В случае неудачи при подъеме крыса была вынуждена выполнить 10 попыток с последним успешным весом без дальнейшего увеличения веса [
15 ].
2.3. Водный лабиринт Морриса
Пространственное обучение и память исследовались с помощью задачи «Водный лабиринт Морриса» [
16]. Круглый резервуар имел диаметр 136 см, высоту 60 см и глубину 30 см, заполненный водой с температурой 24 ± 1 ° C. Лабиринт был разделен на четыре равных квадранта, и каждый квадрант точек высвобождения был назначен на четыре точки: север (N), восток (E), юг (S) и запад (W). Скрытая круглая платформа (диаметром 10 см), погруженная на 1,5 см ниже поверхности воды, располагалась в центре юго-восточного квадранта. Различные места давали визуальные подсказки по лабиринту. Задержка при побеге и пройденное расстояние были измерены для системы слежения. Крысы OLETF и LETO проводили четыре испытания в день в течение пяти дней подряд на платформе, находящейся в одном и том же положении. Мы случайным образом назначали начальные точки каждый день. Мы прекращали испытания либо тогда, когда крысы OLETF и крысы LETO забирались на платформу, либо по истечении 60 с. После подъема на платформу крысам OLETF и LETO позволяли оставаться на платформе в течение 15 с. Когда крыса не находила платформу в течение 60 с, крысу перемещали на платформу и присваивали оценку 60 с. После завершения четырех испытаний крысы OLETF и LETO вернулись в свои клетки с теплой средой. Задержки эвакуации и пройденное расстояние на последней платформе были рассчитаны с использованием компьютеризированной системы видеонаблюдения (Noldus Ethovision Version 5, Wageningen, Нидерланды). Мы определили ежедневные баллы как средние баллы четырех испытаний, выполненных в день. Крысы OLETF и LETO выполнили 60-секундный зондовый след с платформой, удаленной из резервуара по данным. Время, проведенное в целевом квадранте, рассчитывалось с использованием компьютеризированной системы видеонаблюдения (Noldus Ethovision Version 5, Wageningen, Нидерланды).
2.4. Биохимия крови
Вкратце, экспериментальных крыс умерщвляли изофлураном и собирали образцы цельной крови натощак в течение ночи. Кровь экспериментальных крыс получали при помощи левого бокового доступа калибра 20 при пункции сердца. Сыворотка была получена из цельной крови 9 мл в пустых пробирках и позволила крови полностью свернуться. Мы центрифугировали каждую пробирку при 2500 × g в течение 15 мин и переносили сыворотку в пустую пробирку для биохимического анализа. Мы проанализировали уровни глюкозы натощак с помощью глюкометра One Touch II (Life Scan, Johnson & Johnson, Нью-Брансуик, Нью-Джерси, США). Уровни инсулина в сыворотке, общего холестерина и триглицеридов (TG) в сыворотке были получены с помощью иммуноферментного анализа (ELISA; ALPCO, Салем, штат Нью-Хэмпшир, США) и ферментативных наборов (Wako Chemicals USA, Inc., Ричмонд, Вирджиния, США). .
2.5. Анализ клеточного цикла
Кишки крыс получали после умерщвления, когда крыс кормили в течение 8 недель. Клетки кишечного эпителия (IEC) выделяли с использованием Dispase (Sigma-Aldrich, Сент-Луис, Миссури, США) и сетчатого фильтра для клеток 45 мкм, дважды промывали PBS, фиксировали 70% этанолом и окрашивали йодидом пропидия (PI; Invitrogen, Waltham, Massachusetts, USA) для анализа клеточной сортировки по флуоресценции (FACS).
2.6. Окрашивание гематоксилином и эозином (H&E)
Печень, скелетные мышцы, тонкий кишечник и кожные ткани крыс фиксировали в 4% параформальдегиде и заключали в парафиновые блоки. Блоки делали на срезы размером 3 мкм, и срезы окрашивали гематоксилином и эозином (H&E). Морфологический анализ проводился с использованием системы анализатора изображений Leica Qwin, оснащенной микроскопом Leica DMLS.
2.7. Экстракция РНК и синтез кДНК
Реагент TRI от Bio-Fact (Тэджон, Корея) использовали для выделения общей РНК из 30 мг цельной ткани кишечника в соответствии с инструкциями производителя. РНК количественно определяли с помощью Nano Drop (Nano Drop ONE, Thermo Scientific Inc., Уолтем, Массачусетс, США). Для синтеза кДНК использовали RT-Kit (M-MLV, RNase H-, Bio-Fact Co., Daejeon, Korea).
2.8. Количественный КПЦР в реальном времени
Уровни экспрессии мРНК определяли с помощью ПЦР в реальном времени (Agilent Co., Санта-Клара, Калифорния, США) с использованием 2X Master Mix для ПЦР в реальном времени (включая SYBR Green I, Low ROX, Bio-Fact Co.). Анализ данных выполнялся с помощью программы Agilent Aria MX 1.0. Последовательности праймеров для крысиного IL6, TNFα, IL10, BiP и β-актина были следующими; (IL6) fw: TCC TAC CCC AAC TTC CAA TGC TC, rv: TTG GAT GGT CTT GGT CCT TAG CC, (TNFα) fw: GTC GTA GCA AAC CAC CAA G, rv: AGA GAA CCT GGG AGT AGA TAA G, ( IL10) fw: ATT GAA CCA CCC GGC ATC TA, rv: CAA CGA GGT TTT CCA AGG AG и (BiP) fw: AGA AAC TCC GGC GTG AGG TAG A, rv: TTT CTG GAC AGG TTT CAT GGT AG, (β -актин) fw: GGC ACC ACA CTT TCT ACA AT, rv: AGG TCT CAA ACA TGA TCT GG
2.9. Обратная транскрипция-ПЦР
Обратная транскрипция (RT) -PCR выполнялась с использованием 2X Taq Basic PCR Master Mix в соответствии с инструкциями производителя (BioFACT Co., Корея). Образцы загружали в 3% агарозный гель для электрофореза и визуализировали в УФ-свете (AE-9000 E-Graph, ATTO, Tokyo, Japan). Последовательности праймеров крысиного CHOP и гипоксантинфосфорибозилтрансферазы (HPRT) были следующими; (CHOP) fw: GTC TCT GCC TTT CGC CTT TG, rv: AGC TGG AAG CCT GGT ATG AGG A, (HPRT) fw: GCT GAC CTG CTG GAT TAC AT, rv: CCC GTT GAC TGG TCA TTA CA.
2.10. Статистический анализ
Статистический анализ был выполнен с использованием однофакторного дисперсионного анализа для всех данных и с использованием двустороннего дисперсионного анализа с повторными измерениями с последующим апостериорным тестом Тьюки для поведенческих данных. Результаты представлены в виде средних значений ± стандартные ошибки, а статистическая значимость была утверждена выше 0,05 от р -значения
3. Результаты
3.1. Влияние упражнений и добавок куркумина на метаболические осложнения у крыс OLETF
Мы использовали крыс Otsuka Long-Evans Tokushima Fatty (OLETF), которая является хорошо известной моделью сахарного диабета 2 типа (T2DM), и крыс Long-Evans Tokushima Otsuka (LETO) в качестве контроля без диабета. У крыс OLETF, полученных из лаборатории Charles River и Исследовательского института Tokushima, спонтанно развивается диабет с фенотипами полиурии, полидипсии и легкого ожирения [
17 ]. Первоначально мы исследовали метаболизм глюкозы и липидов у крыс OLEFT и LETO, а затем исследовали, оказывают ли упражнения и добавки куркумина положительное влияние на крыс OLETF.
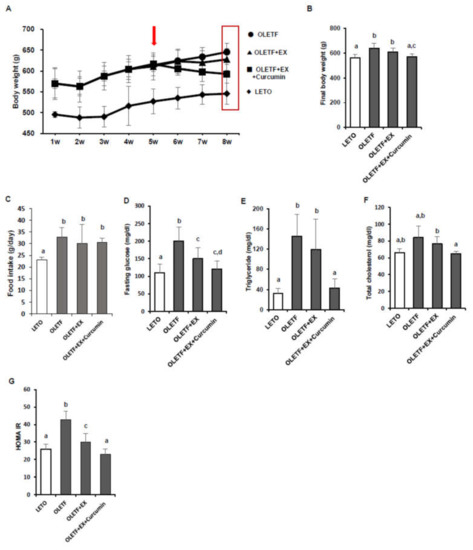
Мы измерили изменения массы тела, уровня глюкозы натощак и показателей HOMA-IR ( A, B). Средняя конечная масса тела была значительно выше для контрольных групп OLETF, чем для контролей LETO ( p <0,05). Средняя масса тела была значительно ниже в группе OLETF + EX + куркумин, чем в контрольной группе OLETF ( p <0,05), тогда как между контрольной группой OLETF и крысами в группе OLETF + EX не наблюдалось значительной разницы в средней массе тела ( А, Б). Среднее потребление пищи (г / день) было значительно выше в контроле OLETF, чем в контроле LETO ( p <0,05), как и ожидалось, и не было значительных различий между контролями OLETF, OLETF + EX и группой OLETF + EX + куркумин ( C). Контроли OLETF имели значительно более высокие уровни глюкозы в крови натощак и HOMA-IR, чем контроли LETO, а уровни глюкозы в крови натощак и HOMA-IR были значительно ниже в группах OLETF + EX и OLETF + EX + куркумин, чем в контрольных группах OLETF, и были ниже. в группе OLETF + EX + куркумин, чем в группе OLETF + EX ( p <0,05; D, E).
Рис. 1. Физические упражнения плюс добавка куркумина уменьшили массу тела и улучшили гомеостаз глюкозы и липидный профиль у крыс OLETF. ( A ) График массы тела (г) в течение восьми недель эксперимента для контроля OLETF и LETO (10 самцов крыс / группа). Стрелка указывает на начальный период исследования интервенционных упражнений с добавлением куркумина. ( B ) Средняя масса тела была значительно ниже в группе OLETF + EX + куркумин, чем в контрольной группе OLETF, и ниже в группе OLETF + EX + куркумин, чем в группе OLETF + EX. ( C ) Потребление пищи (г / день) измеряли два раза в неделю. ( DУровни глюкозы натощак были ниже в группах OLETF + EX + куркумин и OLETF + EX, чем в контрольной группе OLETF, но между этими группами вмешательства не наблюдалось значительной разницы. ( E — G ) Триглицериды сыворотки, общий холестерин и гомеостатическая модель оценки резистентности к инсулину (HOMA-IR) в контроле OLETF и LETO. Результаты представлены как средние значения ± стандартное отклонение (SD). Значимые различия определяли с помощью однофакторного дисперсионного анализа. Разные верхние индексы указывают на значительную разницу; а, б, в, г ( р <0,05). LETO: соответствующая возрасту контрольная группа, OLETF: диабетическая группа, OLETF + EX: диабетическая группа, тренированная физическими упражнениями (для подъема по лестнице с дополнительными весами), OLETF + EX + Куркумин: диабетическая группа, тренируемая физическими упражнениями и получающая куркумин (5 г / кг) / диета) группа.
Чтобы определить, оказывают ли упражнения плюс добавка куркумина положительное влияние на метаболизм липидов у крыс OLETF, были проанализированы уровни триглицеридов в сыворотке и общего холестерина. Контроли OLETF имели значительно более высокие уровни триглицеридов ( p <0,05) и общего холестерина ( p <0,05), чем контроли LETO. Интересно, что триглицериды и общий холестерин были значительно ниже в группе OLETF + EX + куркумин, чем в контрольной группе OLETF, но не наблюдалось значительной разницы между группой OLETF + EX и контролем OLETF ( F, G).
3.2. Влияние упражнений и куркумина на гистопатологию у крыс OLETF
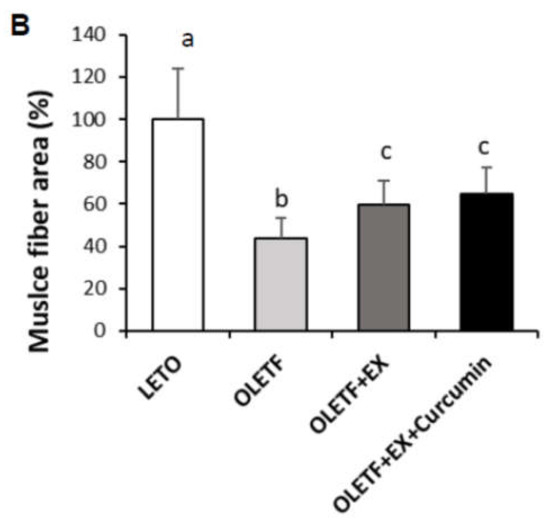
Мы изучили влияние упражнений и добавок куркумина на морфологические изменения в тканях печени и мышц с помощью окрашивания H&E ( ). Печеночные ткани контрольных животных OLETF показали умеренную вакуолизацию, тогда как в контрольных группах LETO этого не наблюдалось. Интересно, что в группе OLETF + EX + куркумин печеночная и мышечная ткани выглядели нормальными без вакуолизации ( A). Ни LETO, ни OLETF контроли не проявляли аномальных характеристик, таких как мышечный фиброз икроножных мышц. Однако размеры мышечных волокон были значительно меньше в контроле OLETF по сравнению с контролем LETO ( p<0,05), а размер мышечных волокон как в группе OLETF + EX, так и в группе OLETF + EX + куркумин был значительно больше в контрольной группе OLETF. В срезах кожи контрольные группы OLETF имели менее развитые слои кожных мышц под гиподермой, чем контроли LETO. Интересно, что эти слои кожных мышц были намного более развиты в группах OLETF + EX и OLETF + EX + куркумин, чем в контрольной группе OLETF.
Рисунок 2. Куркумин снижает связанные с диабетом фенотипы печени, мышц, кожи и тонкого кишечника у диабетических крыс. ( A ) Типичное окрашивание гематоксилином и эозином (H&E) печени, мышц, кожи и тканей тонкого кишечника контрольных групп OLETF и LETO, полученных в конце восьминедельного экспериментального периода. Физические упражнения плюс куркумин изменили морфологию печени, мышечных слоев кожи (указано стрелкой) и ворсинок тонкой кишки (показано стрелкой). ( B) Количественные данные размера мышечных волокон экспериментальных крыс. Площадь поперечного сечения мышечного волокна измеряли для всех мышечных волокон пяти изображений, выбранных для каждого животного. В данных была рассчитана средняя площадь мышечных волокон и представлена как среднее значение площади всех мышечных волокон на пяти изображениях. Существенные различия были определены с помощью однофакторного дисперсионного анализа с последующим апостериорным тестом Турции. Различные верхние индексы указывают на значительную разницу ( p <0,05). Контроль LETO: контрольная группа соответствующего возраста, контрольная группа OLETF: диабетическая группа, OLETF + EX: диабетическая группа тренировок, OLETF + EX + куркумин: тренировка диабетиков с упражнениями плюс куркумин (5 г / кг в рационе).
Считается, что СД2 сопровождается хроническим воспалением, и даже легкое, но хроническое воспаление может нарушить эпителий тонкой кишки и укорачивать ворсинки. Тонкий кишечник контрольной группы OLETF показал относительно короткие и поврежденные ворсинки по сравнению с контрольной группой LETO, что позволяет предположить, что у крыс OLETF наблюдается хроническое воспаление в тонкой кишке. Таким образом, это наблюдение показало. Кроме того, инфильтрирующие моноциты наблюдались в ворсинах тонкой кишки, как показано стрелками, что обычно наблюдается при воспалительных состояниях, таких как колит. На фотографии, увеличенной в 40 раз, показаны неправильно выровненные инфильтрованные моноциты в ворсинки в группе OLETF. Интересно, что эти явления не наблюдались в группе OLETF + EX + куркумин (см. A, как показано стрелками), тогда как группа OLETF + EX показала менее интенсивную инфильтрацию ворсинок моноцитами по сравнению с группой OLETF, хотя ворсинки были нормальной длины.
3.3. Влияние упражнений и куркумина на стресс ER в тканях тонкого кишечника крыс OLETF
Поскольку кишечник является первым барьером от инородного вещества в организме и местом, где куркумин всасывается, мы решили исследовать тонкий кишечник, чтобы определить, влияет ли куркумин на первое место в организме в отношении вызванного диабетом воспаления и стресса ER. Мы исследовали воспалительные цитокины и уровни маркеров стресса ER в тонком кишечнике по регионам. Как показано на A, двенадцатиперстная кишка контрольных OLETF показала повышающую регуляцию провоспалительных цитокинов (IL6 и TNFα) и понижающую регуляцию противовоспалительного IL10. Удивительно, но упражнения сами по себе не устраняли эти эффекты, но упражнения плюс куркумин значительно снижали эти диабетические эффекты, предполагая, что упражнения плюс куркумин снижали провоспалительную реакцию, связанную с диабетом, и усиливали противовоспалительную реакцию. Кроме того, экспрессия маркера стресса ER BiP ( 3А ) и маркера индуцированного стрессом апоптоза, CHOP ( 3С) была значительно ниже в группе OLETF + EX + куркумин, чем в контрольной группе OLETF и группе OLETF + EX. группа.
Рисунок 3. Добавка куркумина с упражнениями подавляла провоспалительные цитокины и стресс ER в тонком кишечнике. ( A — C ) Собирали всю двенадцатиперстную кишку ( A , C ) и тощую кишку ( B ), экстрагировали РНК и синтезировали кДНК для ОТ-ПЦР, нацеленной на маркер апоптоза C / EBP гомологичный белок (CHOP) ( C ) и ПЦР в реальном времени, нацеленная на провоспалительные цитокины (IL6 и TNFα (провоспалительные цитокины), IL10 (противовоспалительный цитокин)) и маркер стресса ER BiP ( A , B ). Гипоксантинфосфорибозилтрансфераза (HPRT) и β-актин использовались в качестве контроля загрузки для ОТ-ПЦР и ПЦР в реальном времени соответственно. (D ) В контрольной группе OLETF куркумин в сочетании с упражнениями спасали фазу G0 / G1 клеточного цикла в изолированных клетках тонкого кишечника. Изолированные клетки тонкой кишки фиксировали 70% этанолом и окрашивали йодидом пропидия (PI). Проточную цитометрию использовали для количественного определения клеточных фаз (% от оси Y). Существенные различия были определены с использованием однофакторного дисперсионного анализа с последующим апостериорным тестом Турции. Различные верхние индексы указывают на значительную разницу ( p <0,05). Результаты представлены в виде средних значений ± стандартные ошибки. Контрольная группа LETO: соответствующая возрасту контрольная группа, контрольная группа OLETF: диабетическая группа, группа OLETF + EX: группа диабетиков, тренированная физическими упражнениями, OLETF + EX + куркумин: группа диабетиков, тренированная физическими упражнениями, получавшая куркумин (5 г / кг в рационе).
Аналогичная картина наблюдалась и в тощей кишке. Как показано на 3B, уровни IL6 и TNFα были значительно выше в контроле OLETF, чем в контроле LETO, но этот эффект значительно уменьшался при использовании упражнений плюс куркумин в большей степени, чем при использовании только упражнений. Кроме того, BiP был значительно ниже в группе OLETF + EX + куркумин, чем в группе OLETF + EX и в контрольной группе OLETF. Эти данные показывают, что упражнения в сочетании с добавлением куркумина оказали заметное влияние на воспаление, связанное с диабетом, и стресс ER, но эти упражнения сами по себе не имели такого заметного эффекта.
Кроме того, мы исследовали, различается ли клеточный цикл клеток тонкой кишки в контроле OLETF и LETO. Мы наблюдали, что фаза G0 / G1 клеточного цикла была ослаблена в группе OLETF + EX и контроле OLETF по сравнению с контролем LETO ( D), но что фаза G0 / G1 была выше в группе OLETF + EX + куркумин, чем в группе OLETF + EX и в контроле OLETF ( p <0,05). Эти наблюдения предполагают, что сниженные уровни мРНК CHOP и уровни мРНК BiP, вызванные упражнениями плюс добавление куркумина, могут увеличивать процент клеточного цикла G0 / G1 на стадии выживания или пролиферации клеток и, таким образом, улучшать когнитивные функции у крыс OLETF.
3.4. Влияние упражнений и добавок куркумина на обучаемость и дефицит памяти у крыс OLETF
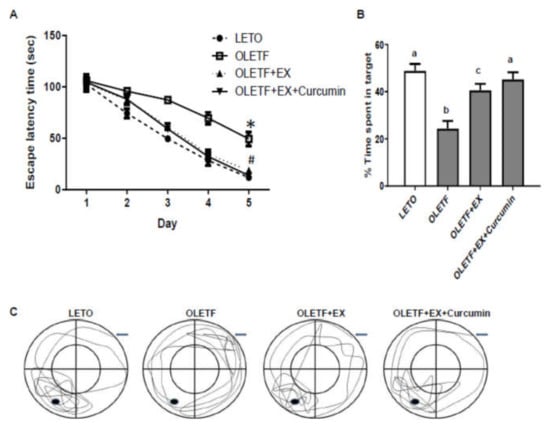
Когнитивные функции оценивались с помощью теста водного лабиринта Морриса. Средняя латентность побега уменьшилась в течение последовательных пяти дней периода обучения во всех группах ( 3А). Мы обнаружили, что средняя латентность побега была больше для OLETF, чем для контроля LETO (49,3 с против 11,8 с у крыс OLETF по сравнению с крысами LETO в системе слежения, p <0,05), но значительно меньше в группе OLETF + EX + куркумин (13,8 с). чем в группе OLETF + EX (18,8 с) и в контроле OLETF (49,4 с) ( A).
Рисунок 4. Куркумин в сочетании с упражнениями улучшил обучение и сохранение памяти у крыс OLETF в тесте в водном лабиринте Морриса. ( A ) Задержка побега измерялась с установленной платформой. По четыре испытания в день экспериментальные крысы выполняли для получения данных в течение пяти дней подряд. * p <0,05 по сравнению с контрольными крысами LETO, # p <0,05 по сравнению с крысами OLETF в контроле. ( B — C ) Влияние упражнений и куркумина на сохранение памяти в испытании с зондом. Время, проведенное в целевом квадранте в испытании зонда, было рассчитано на шестой день ( B ). Типичная диаграмма трекинга по длине плавательной дорожки. Данные были определены с помощью системы видеонаблюдения ( CКонтрольная группа LETO: соответствующая возрасту контрольная группа, контрольная группа OLETF: диабетическая группа, OLETF + EX группа: диабетическая группа тренировок, OLETF + EX + куркумин группа: диабетическая группа тренировок и куркумина (диета 5 г / кг). Результаты представлены в виде средних значений ± стандартные ошибки. Статистический анализ проводился с использованием однофакторного дисперсионного анализа для пробного исследования и с использованием двустороннего дисперсионного анализа с повторными измерениями с последующим апостериорным тестом Турции для поведенческих данных. Различные верхние индексы указывают на значительную разницу ( p <0,05).
Чтобы оценить сохранение памяти, мы провели испытание зонда через два дня после последнего теста в водном лабиринте Морриса. Крыс допускали к плаванию в течение 60 с в бассейне, в котором они тренировались, но с убранной платформой для спасения. Процент времени плавания, проведенного в пределах целевого квадранта у контрольных групп OLETF, был меньше, чем у контрольных групп LETO (24,4% против 48,9% крыс OLETF по сравнению с крысами LETO в системе слежения, p <0,05) и крысами в OLETF +. EX (40,6%) и OLETF + EX + куркумин (45,2%) и был значительно меньше в группе OLETF + EX, чем в группе OLETF + EX + куркумин ( B, C). Эти результаты показывают, что упражнения плюс куркумин могут улучшить когнитивные функции.
4. Обсуждение
В этом исследовании мы предположили, что комбинация упражнений и куркумина может синергетически ослаблять воспалительную реакцию при диабете. Наши результаты показывают, что упражнения вместе с добавкой куркумина ослабляли диабетические фенотипы у крыс OLETF, что определялось измерениями массы тела, глюкозы, инсулина, триглицеридов и общего холестерина. В частности, мы обнаружили, что упражнения с куркумином повышают чувствительность к инсулину, как определено индексами HOMA-IR, и снижают уровень глюкозы натощак у крыс OLETF.
Воспаление — это фундаментальный биологический процесс, который восстанавливает гомеостаз тканей с помощью различных механизмов восстановления и влияет на многие острые и хронические патологические состояния. Однако правильная регуляция воспалительных механизмов важна для предотвращения неконтролируемого усиления начальных воспалительных реакций и перехода от восстановления тканей к побочным повреждениям и развитию заболеваний [
18 ]. Следовательно, легкое хроническое воспаление является распространенным фенотипом метаболического синдрома, включая СД2. Было показано, что стресс ER играет важную роль, а также является обычным явлением между воспалением и СД2, а также многими другими метаболическими нарушениями [
19]. Хотя причины, приводящие к стрессу ER, до сих пор не определены, предполагается, что причиной является повышение циркулирующих провоспалительных цитокинов, таких как TNFα и IL6, в этих метаболических условиях [
8 ]. При T2DM усиление передачи сигналов IRE1α активирует путь NF-kB путем фосфорилирования N-концевой киназы 1 c-jun (JNK), хорошо известных модуляторов воспаления, способствующих экспрессии генов, и активности белков, участвующих в регуляции воспалительной реакции. Это также приводит к нарушению передачи сигналов инсулина и активации нескольких молекул в скелетных мышцах, жировой ткани и печени [
8 ]. Сообщалось, что нарушение передачи сигналов инсулина в результате стресса в ER приводит к системной инсулинорезистентности, гипергликемии и гиперинсулинемии [
7]. В исследовании наши результаты с использованием крыс OLETF, естественно индуцированной модели СД2, подтвердили повышенную инсулинорезистентность, представленную как HOMA-IR на контрольной группе OLETF, и что этот фенотип был улучшен с помощью комбинированной терапии упражнений и добавок куркумина.
Диабет является примером патологического нейродегенеративного процесса, связанного с хроническими воспалительными изменениями. Известно, что основные механизмы диабета и когнитивного дефицита включают повышенные уровни фосфорилированного тау-белка (p-Tau) в спинномозговой жидкости и длительный некодирующий РНК-индуцированный апоптоз нейронов гиппокампа у пожилых диабетических животных и пациентов [
20 ,
21 ]. Несколько исследований показали, что упражнения уменьшают воспалительную реакцию мозга при диабете [
22 ,
23 ,
24 ,
25 ,
26]. Однако другие исследования на людях показали, что изменения одного фактора образа жизни, такие как изменения физической активности, в меньшей степени влияют на когнитивные функции при нейродегенеративных заболеваниях. Куркумин является хорошо известным природным антиоксидантом, который снижает воспалительную реакцию при диабете через сигнальные пути Nrf2 и PI3K / AKT [
27 ,
28 ,
29 ,
30 ,
31 ]. Данные показывают, что куркумин может улучшать патологии, связанные с амилоидом-β, и восстанавливать патологии, связанные с p-тау-белком, на животных моделях [
32 ,
33 ]. Также было показано, что куркумин снижает экспрессию гена рецептора инсулина и субстрата-1 рецептора инсулина в гиппокампе [
34 ,
35 ,
36 ]. Соответственно, наши результаты показывают, что комбинация упражнений и куркумина синергетически улучшает когнитивные функции у крыс OLETF или воспалительную реакцию в кишечнике OLETF.
В модели мышей со сниженным стрессом ER, специфической для мозга, тяжелая резистентность к лептину значительно увеличивала ожирение на диете с высоким содержанием жиров, что предполагает, что стресс ER может участвовать в регулировке чувствительности к инсулину в мозге [
37 ]. Во многих предыдущих исследованиях мы предположили, что стресс ER, наблюдаемый в кишечнике в нашем исследовании, также происходит в мозге, поскольку стресс ER возникает в различных тканях системно, и постулировали, что подавление стресса ER кишечника с помощью упражнений и добавок куркумина произойдет в ткань головного мозга, предполагая, что косвенное подавление стресса ER в головном мозге может улучшить когнитивные функции мозга [
18 ,
19 ,
38 ,
39 ,
40 ,
41].
Новые доклинические данные показали, что двунаправленная передача сигналов между желудочно-кишечным трактом (ЖКТ) и мозгом, так называемая ось кишечник-мозг, играет важную роль как в метаболизме, так и в поведении хозяина [
42 ]. Имеющиеся данные свидетельствуют о том, что микробное разнообразие снижается при нарушении регуляции метаболизма [
43 ,
44 ,
45 ]. Обнаружено, что микробные метаболиты короткоцепочечных жирных кислот оказывают множество физиологических эффектов, включая энергетический гомеостаз, за счет регуляции гормонов ЖКТ, таких как холецистокинин, глюкагоноподобный пептид 1 (GLP1) и лептин [
46]. Более того, кишечные микробиомы вызывают изменения в секреции кишечных гормонов и иммунном ответе, а также в барьере слизистой оболочки кишечника, контроле гематоэнцефалического барьера и создании и разложении нейромедиатора, таким образом, тесно взаимодействуя с центральной нервной системой, что влияет на способность мозга развивать когнитивное поведение. Корреляции между микробиомом кишечника и так называемой осью мозг-кишечник интенсивно изучаются при неврологических заболеваниях. Одно исследование показало, что потеря толл-подобного рецептора (TLR) 2, одного из датчиков врожденного иммунитета, у традиционных мышей приводит к напоминающему фенотип метаболического синдрома, характеризующемуся четким различием микробиоты кишечника, что вызывает резистентность к инсулину. и воспаление, связанное со стрессом ER,
47 ]. Эти данные подчеркнули роль микробиоты в сложной сети молекулярных и клеточных взаимодействий, которые связывают генотип с фенотипом и имеют потенциальные последствия для широкого спектра распространенных заболеваний человека, включая диабет, и даже других иммунологических нарушений [
43 ].
Однако у этого исследования было несколько ограничений. Во-первых, исследование было сосредоточено на влиянии упражнений крыс OLETF на когнитивную функцию, которое не обеспечивает экспериментальной группе крыс OLETF только куркумин. Тем не менее, упражнения являются основным фактором образа жизни взрослых в отношении когнитивной функции, что также было включено в исследование для изучения влияния куркумина. Во-вторых, мы не предоставили данные гистологии или ER стресса в головном мозге. Однако наша лаборатория ранее сообщала, что у крыс OLEF с упражнениями в гипоталамусе наблюдалось значительное повышение уровней мРНК GLP-1 (основного инкретинового гормона), переносчика глюкозы 2 (GLUT2) и супероксиддисмутазы 1 (SOD1, антиоксидантный фермент) по сравнению с Крысы OLETF в тканях мозга [
48]. Действительно, как упоминалось выше, мы обнаружили, что упражнения в сочетании с куркумином устраняют стрессовую реакцию ER в тканях кишечника контрольной группы OLETF и повышают чувствительность к инсулину в системе крови. Кроме того, крысы в группе OLETF + EX + куркумин продемонстрировали лучшие навыки обучения и память мозга в тесте в водном лабиринте Морриса, который предполагает, что упражнения плюс куркумин регулируют чувствительность к инсулину во всем организме, снижая стресс ER и повышая резистентность к лептину. В результате уровень инсулина регулируется, стресс ER, вызванный высоким уровнем глюкозы в крови, снижается, а апоптоз, вызванный стрессом ER, подавляется в системе крови, а когнитивные функции улучшаются у крыс OLETF. Необходимы дальнейшие исследования для подтверждения результатов в различных тканях, включая мозг, с точки зрения воспаления и стресса ER, чтобы уменьшить симптомы диабетической болезни.
В качестве третьего ограничения куркумин, как биоактивное соединение, растворим в этаноле, щелочи, уксусной кислоте и других органических растворителях и плохо всасывается в кишечнике. В одном исследовании сообщалось о всасывании 400 мг куркумина, введенного перорально крысам, и было обнаружено в 5 мг / л в воротной вене печени через 24 часа после пероральной инъекции [
49 ,
50 ].
Однако в этом исследовании мы наблюдали, что крысы, получавшие куркумин в количестве 5 г на рацион (кг), улучшали потерю веса, гомеостаз глюкозы и липидный профиль. Несмотря на эти ограничения, имеющиеся данные позволяют сделать вывод, что сочетание упражнений и куркумина улучшает когнитивные явления, связанные с диабетом, у крыс.
Вклад авторов
Концептуализация, EP, JAC и JHY; методология, SHP; ресурсы, EP и JAC; курирование данных, JC и J.-OK; Подготовка оригинального черновика, JAC и EP; финансирование привлечения, EP и JAC Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование было поддержано Программой фундаментальных научных исследований через Национальный исследовательский фонд Кореи, финансируемой Министерством образования: NRF-2019R1H1A2039746 (EP) и NRF-2019R1A2B5B01070133 (JAC).
Благодарности
Мы благодарим Хёну Ли, Хэлим Пак, Хе-Чон Хван и Джу-Гён Юн за их помощь во всех экспериментах на животных.
Конфликт интересов
Авторы не заявляют о конфликте интересов.
Ссылки
- Биссельс, ГДж; Реймер, Ю. Д. Изменения мозга, лежащие в основе когнитивной дисфункции при диабете: чему мы можем научиться с помощью МРТ? Диабет 2014 , 63 , 2244–2252. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Биссельс, ГДж; Страчан, MWJ; Visseren, FL; Каппелле, LJ; А. Уитмер, Р. Деменция и снижение когнитивных функций при диабете 2 типа и предиабетических стадиях: на пути к целевым мероприятиям. Ланцет Диабет Эндокринол. 2014 , 2 , 246–255. [ Google Scholar ] [ CrossRef ]
- Riederer, P .; Корчин, А.Д .; Али, СС; Bajenaru, O .; Choi, MS; Чопп, М .; Дерманович-Доброта, В .; Grünblatt, E .; Jellinger, KA; Камаль, Массачусетс; и другие. Диабетический мозг и познание. J. Neural Transm. 2017 , 124 , 1431–1454. [ Google Scholar ] [ CrossRef ]
- Чжао, Р.Р .; О’Салливан, AJ; Сингх, MAF. Упражнения или физическая активность и когнитивные функции у взрослых с диабетом 2 типа, инсулинорезистентностью или нарушенной толерантностью к глюкозе: систематический обзор. Евро. Rev. Aging Phys. Закон. 2018 , 15 , 1. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Хардинг, HP; Zhang, Y .; Ron, D. Трансляция и фолдинг белка связаны с киназой, резидентной в эндоплазматическом ретикулуме. Nature 1999 , 397 , 271–274. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hwang, J .; Ци, Л. Контроль качества в эндоплазматической сети: перекрестные помехи между путями ERAD и UPR. Trends Biochem. Sci. 2018 , 43 , 593–605. [ Google Scholar ] [ CrossRef ]
- Ozcan, U .; Yilmaz, E .; Ozcan, L .; Furuhashi, M .; Vaillancourt, E .; Смит, РО; Гёргюн, Чехия; Хотамислигил, Г.С. Химические шапероны уменьшают стресс ER и восстанавливают гомеостаз глюкозы на мышиной модели диабета 2 типа. Science 2006 , 313 , 1137–1140. [ Google Scholar ] [ CrossRef ]
- Хотамислигил, Г.С. Стресс эндоплазматического ретикулума и воспалительная основа метаболических заболеваний. Cell 2010 , 140 , 900–917. [ Google Scholar ] [ CrossRef ]
- Chen, R .; Shi, J .; Инь, Q .; Li, X .; Sheng, Y .; Han, J .; Zhuang, P .; Чжан Ю. Морфологические и патологические характеристики мозга при диабетической энцефалопатии. J. Alzheimers Dis. 2018 , 65 , 15–28. [ Google Scholar ] [ CrossRef ]
- Чжан, X .; Xu, L .; Он, Д .; Линг, С. Эндоплазматический ретикулум, вызванный стрессом, апоптоз нейронов гиппокампа, вовлеченный в диабетическое когнитивное нарушение. BioMed Res. Int. 2013 , 2013 , 1–9. [ Google Scholar ] [ CrossRef ]
- Чо, JA; Park, E. Куркумин использует путь противовоспалительной реакции для защиты кишечника от бактериальной инвазии. Nutr. Res. Практик. 2015 , 9 , 117–122. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Чой, Ю.Е .; Park, E. Куркумин повышает чувствительность ингибитора поли (АДФ-рибозы) полимеразы к химиотерапии в клетках рака груди. J. Nutr. Biochem. 2015 , 26 , 1442–1447. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Буглиани, М .; Mossuto, S .; Grano, F .; Сулейман, М .; Marselli, L .; Boggi, U .; De Simone, P .; Эйзирик, DL; Cnop, M .; Marchetti, P .; и другие. Модуляция аутофагии влияет на функцию и выживаемость бета-клеток поджелудочной железы человека в условиях стресса эндоплазматической сети и при диабете 2 типа. Фронт. Эндокринол. 2019 , 10 , 52. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kawano, K .; Hirashima, T .; Mori, S .; Saitoh, Y .; Куросуми, М .; Натори, Т. Спонтанная длительная гипергликемия у крыс с диабетическими осложнениями: штамм Оцука Лонг-Эванс Токусима жирный (OLETF). Диабет 1992 , 41 , 1422–1428. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kim, H.-J .; Сонг, У. Тренировка с отягощениями увеличивает уровень фактора роста фибробластов-21 и уровень иризина в скелетных мышцах крыс с диабетом Цукера с ожирением. J. Exerc. Nutr. Biochem. 2017 , 21 , 50–54. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Vorhees, CV; Уильямс, М. Т. Водный лабиринт Морриса: процедуры для оценки пространственных и связанных форм обучения и памяти. Nat. Protoc. 2006 , 1 , 848–858. [ Google Scholar ] [ CrossRef ]
- Kawano, K .; Hirashima, T .; Mori, S .; Натори, Т. OLETF (Otsuka Long-Evans Tokushima Fatty) крыса: новая линия крыс с NIDDM. Diabetes Res. Clin. Практик. 1994 , 24 , S317 – S320. [ Google Scholar ] [ CrossRef ]
- Sims-Robinson, C .; Zhao, S .; Hur, J .; Фельдман, Е. Л. Стресс эндоплазматического ретикулума центральной нервной системы на мышиной модели диабета 2 типа. Диабетол. 2012 , 55 , 2276–2284. [ Google Scholar ] [ CrossRef ]
- Чжан, X .; Zhang, G .; Zhang, H .; Карин, М .; Bai, H .; Цай, Д. Гипоталамический IKKβ / NF-κB и ER стресс связывают переедание с энергетическим дисбалансом и ожирением. Cell 2008 , 135 , 61–73. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Yu, J .; Li, C .; Че, Л .; Zhao, Y .; Guo, Y. Подавление длинной некодирующей РНК H19 спасает нейроны гиппокампа от апоптоза и окислительного стресса, ингибируя метилирование IGF2 у мышей с сахарным диабетом, индуцированным стрептозотоцином. J. Cell. Physiol. 2018 , 234 , 10655–10670. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Moran, C .; Beare, R .; Phan, TG; Брюс, Д.Г.; Каллисая, М .; Срикант, В.К. Сахарный диабет 2 типа и биомаркеры нейродегенерации. Неврология 2015 , 85 , 1123–1130. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Белотто, MF; Magdalon, J .; Родригес, HG; Виноло, МАР; Curi, R .; Пифон-Кури, ТС; Хатанака, Э. Умеренные упражнения улучшают функцию лейкоцитов и уменьшают воспаление при диабете. Clin. Exp. Иммунол. 2010 , 162 , 237–243. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Karstoft, K .; Педерсен, Б.К. Упражнения и диабет 2 типа: акцент на метаболизм и воспаление. Иммунол. Cell Boil. 2015 , 94 , 146–150. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- De Lemos, ET; Пинто, РМ; Oliveira, J .; Garrido, P .; Sereno, J .; Mascarenhas-Melo, F .; Пинейро, JP; Тейшейра, Ф .; Рейс, Ф. Дифференциальные эффекты острых (ослабляющих) и хронических (тренировочных) упражнений на воспаление и статус окислительного стресса на животной модели сахарного диабета 2 типа. Mediat. Воспаление. 2011 , 2011 , 1–8. [ Google Scholar ] [ CrossRef ]
- Waddington, G. Интенсивность упражнений и воспаление при диабете 2 типа. J. Sci. Med. Спорт 2017 , 20 , 885. [ Google Scholar ] [ CrossRef ]
- Zoppini, G .; Targher, G .; Zamboni, C .; Вентури, C .; Cacciatori, V .; Moghetti, P .; Muggeo, M. Влияние тренировок средней интенсивности на биомаркеры воспаления и эндотелиальной дисфункции у пожилых пациентов с диабетом 2 типа. Nutr. Метаб. Кардиоваск. Dis. 2006 , 16 , 543–549. [ Google Scholar ] [ CrossRef ]
- Biswas, S .; Chen, S .; Liang, G .; Feng, B .; Cai, L .; Хан, З.А.; Чакрабарти, С. Аналоги куркумина снижают показатели стресса и воспаления в экспериментальных моделях диабета. Фронт. Эндокринол. 2019 , 10 , 887. [ Google Scholar ] [ CrossRef ]
- Ghosh, S .; Banerjee, S .; Sil, PC Благоприятная роль куркумина при воспалении, диабете и нейродегенеративных заболеваниях: недавнее обновление. Food Chem. Toxicol. 2015 , 83 , 111–124. [ Google Scholar ] [ CrossRef ]
- Рашид, К .; Chowdhury, S .; Ghosh, S .; Sil, PC Куркумин ослабляет вызванное окислительным стрессом воспаление, опосредованное NFκB, и зависимый от эндоплазматического ретикулума апоптоз спленоцитов при диабете. Biochem. Pharmacol. 2017 , 143 , 140–155. [ Google Scholar ] [ CrossRef ]
- Weisberg, S .; Leibel, R .; Торториелло, Д.В. Диетический куркумин значительно снижает воспаление, связанное с ожирением, и диабет на мышиных моделях диабета. Эндокринология 2008 , 149 , 3549–3558. [ Google Scholar ] [ CrossRef ]
- Ян, З .; Dai, Y .; Fu, H .; Zheng, Y .; Bao, D .; Инь, Й .; Chen, Q .; Не, X .; Hao, Q .; Hou, D.-R .; и другие. Куркумин оказывает защитное действие против преждевременной недостаточности яичников у мышей. J. Mol. Эндокринол. 2018 , 60 , 261–271. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Привет.; Wang, P .; Wei, P .; Feng, H .; Ren, Y .; Yang, J .; Rao, Y .; Shi, J .; Тиан, Дж. Влияние куркумина на синапсы у мышей APPswe / PS1dE9. Int. J. Immunopathol. Pharmacol. 2016 , 29 , 217–225. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Ma, Q.-L .; Zuo, X .; Ян, Ф .; Убеда, О. Дж .; Гант, диджей; Алавердян, М .; Teng, E .; Hu, S .; Chen, P.-P .; Maiti, P .; и другие. Куркумин подавляет растворимые димеры тау-белка и корректирует молекулярные шапероны, синаптические и поведенческие дефициты у старых трансгенных мышей человека. J. Boil. Chem. 2012 , 288 , 4056–4065. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Feng, H.-L .; Dang, H.-Z .; Fan, H .; Chen, X.-P .; Рао, Y.-X .; Ren, Y .; Yang, J.-D .; Shi, J .; Wang, P .; Тиан, Ж.-З. Куркумин улучшает сигнальный путь инсулина в мозге трансгенных мышей с болезнью Альцгеймера. Int. J. Immunopathol. Pharmacol. 2016 , 29 , 734–741. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zhang, L .; Kong, X.-J .; Wang, Z.-Q .; Xu, F.-S .; Чжу, Ю.-Т. Исследование нейропротекторного действия куркумина на мозг крыс с диабетом. J. Nutr. Исцелить. Старение 2016 , 20 , 835–840. [ Google Scholar ] [ CrossRef ]
- Wang, P .; Вс, Ц .; Feng, H .; Чен, X .; Dong, Y .; Rao, Y .; Ren, Y .; Yang, J .; Shi, J .; Tian, J .; и другие. Куркумин регулирует пути инсулина и метаболизм глюкозы в мозге мышей APPswe / PS1dE9. Int. J. Immunopathol. Pharmacol. 2017 , 30 , 25–43. [ Google Scholar ] [ CrossRef ]
- Ozcan, L .; Эргин, АС; Лу, А .; Chung, J .; Sarkar, S .; Nie, D .; Майерс, MG; Озкан, У. Стресс эндоплазматического ретикулума играет центральную роль в развитии резистентности к лептину. Cell Metab. 2009 , 9 , 35–51. [ Google Scholar ] [ CrossRef ]
- Бускетс, О .; Eritja, À .; Лопес, Б.М.; Ettcheto, M .; Manzine, P .; Кастро-Торрес, РД; Verdaguer, E .; Olloquequi, J .; Vázquez-Carrera, M .; Auladell, C .; и другие. Роль N-концевой киназы 2 мозга c-Jun в контроле рецептора инсулина и ее связь с когнитивными функциями в доклинической модели диеты с высоким содержанием жиров. J. Neurochem. 2019 , 149 , 255–268. [ Google Scholar ] [ CrossRef ]
- Kong, F.-J .; Торговый центр.; Guo, J.-J .; Xu, L.-H .; Li, Y .; Ку, S. Путь стресса / аутофагии эндоплазматического ретикулума участвует в индуцированном диабетом апоптозе нейронов и снижении когнитивных функций у мышей. Clin. Sci. 2018 , 132 , 111–125. [ Google Scholar ] [ CrossRef ]
- Muriach, M .; Флорес-Беллвер, М .; Romero, FJ; Барсия, Дж. М. Диабет и мозг: окислительный стресс, воспаление и аутофагия. Oxidative Med. Cell. Longev. 2014 , 2014 , 1–9. [ Google Scholar ] [ CrossRef ]
- Zhao, Y .; Ян, Ю .; Zhao, Z .; Li, S .; Инь, Дж. Динамические изменения маркеров пути стресса эндоплазматического ретикулума GRP78 и CHOP в гиппокампе мышей с диабетом. Brain Res. Бык. 2015 , 111 , 27–35. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Cryan, JF; Динан, Т.Г. Изменяющие сознание микроорганизмы: влияние микробиоты кишечника на мозг и поведение. Nat. Rev. Neurosci. 2012 , 13 , 701–712. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Де Клерк, Северная Каролина; Фриссен, Миннесота; Groen, AK; Nieuwdorp, M. Микробиота кишечника и ось кишечника и мозга. Психосом. Med. 2017 , 79 , 874–879. [ Google Scholar ] [ CrossRef ]
- Arora, T .; Бэкхед, Ф. Микробиота кишечника и метаболические заболевания: текущее понимание и перспективы на будущее. J. Intern. Med. 2016 , 280 , 339–349. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Ianiro, G .; Bibbò, S .; Антонио, G .; Каммарота, Г. Терапевтическая модуляция микробиоты кишечника: текущие клинические применения и перспективы на будущее. Curr. Drug Targets 2014 , 15 , 762–770. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Yadav, H .; Lee, J.-H .; Lloyd, J .; Walter, P .; Рэйн, С.Г. Благоприятные метаболические эффекты пробиотика через секрецию гормона GLP-1, вызванную бутиратом. J. Boil. Chem. 2013 , 288 , 25088–25097. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Карицилли, AM; Пикарди, ПК; Де Абреу, LL; Ueno, M .; Prada, PO; Веревка, ER; Hirabara, SM; Castoldi, A .; Де Мораэш-Виейра, PMM; Камара, н. и другие. Микробиота кишечника является ключевым модулятором инсулинорезистентности у мышей с нокаутом TLR 2. PLoS Кипятить. 2011 , 9 , e1001212. [ Google Scholar ] [ CrossRef ]
- Парк, Ш; Юн, JH; Seo, DY; Ким, TN; Ko, J .; Хан, Дж. Тренировка с отягощениями снижает потерю эндогенного рецептора GLP-1 в гипоталамусе крыс с диабетом 2 типа. Int. J. Environ. Res. Public Health 2019 , 16 , 830. [ Google Scholar ] [ CrossRef ]
- Равиндранатх, V .; Чандрасекхара, Н. Поглощение и распределение куркумина в тканях у крыс. Toxicol. 1980 , 16 , 259–265. [ Google Scholar ] [ CrossRef ]
- Xue, M .; Cheng, Y .; Xu, L .; Чжан, Л. Исследование характеристик абсорбции куркумина в кишечнике in vivo и in vitro. J. Appl. Pharm. 2017 , 9 . [ Google Scholar ] [ CrossRef ]
© 2020 Авторы. Лицензиат MDPI, Базель, Швейцария. Эта статья — статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution (CC BY) (http://creativecommons.org/licenses/by/4.0/ )




Без комментариев